Морфология имаго (терминология)
В комплексе признаков, по которым имаго Rhopalocera отличаются от прочих чешуекрылых, наиболее заметны: булавовидные или головчатые усики, отсутствие простых глазков, полная редукция френатного сцепочного аппарата крыльев, крылья в покое складываются за спиной, уменьшение числа жилок крыльев за счет полного или частичного слияния и редукции - особенно характерно наличие на заднем крыле двух или даже одной анальной жилок, наличие яркого пигментного или структурного рисунка на обеих поверхностях крыльев, значительное упрощение генитального аппарата, активность в дневное время.
Тело булавоусого чешуекрылого состоит из 3 хорошо обособленных отделов: головки (caput), груди (thorax) и брюшка (abdomen).
Голова (рис. 5) округлая (Papilionoidea) или поперечная (Hesperioidea), малоподвижная, гипогнатная (ротовые части расположены снизу). По бокам головной капсулы, занимая большую часть ее поверхности, располагаются крупные полусферические глаза (oculi), состоящие из большого числа фасеток, или омматидиев (ommatidia). Глазная кутикула у некоторых видов снабжена более или менее длинными прямостоящими волосками, расположенными между фасетками (волосистые глаза); у большинства дневных бабочек глаза голые. У верхнепереднего края глаз, в сочленовных ямках лобно-теменного шва, расположены основания антенн или усиков (antennae), состоящих из большого числа члеников, покрытых плотно прилегающими чешуями (рис. 6). В зависимости от характера изменения диаметра члеников в направлении от основания к вершине различают булавовидные, головчатые и веретеновидные усики; у некоторых Hesperiidae утонченная вершина веретеновидных усиков бывает более или менее резко изогнутой (крючковидные усики). Ротовой аппарат у всех булавоусых чешуекрылых представлен длинным, в состоянии покоя свернутым хоботком (proboscis), состоящим из пары слабо сочленованных жевательных лопастей (galea). Все дневные бабочки питаются нектаром цветков и растворами органических веществ. В знойный день их можно видеть у мелких водоемов и на увлажненных участках почвы, где они пьют воду, некоторые виды питаются соком, вытекающим из раненых стволов деревьев, экскрементами, жидкостью трупов и др. По бокам хоботка расположена пара 3-членистых губных щупиков (palpi labiales), торчащих вперед или прилегающих к головной капсуле спереди; у Libythea губные щупики гипертрофированы, их длина намного превосходит наибольший размер головы и даже груди.
Грудной отдел представлен тремя хорошо склеротизованными сегментами: передне-, средне- и заднегрудным. Вентральная часть каждого сегмента несет пару ног, к дорсальной части средне- и заднегруди прикрепляются соответственно передние и задние крылья. Свободная часть ноги состоит из вертлуга (trochanter), бедра (femur), голени (tibia) и членистой лапки (tarsus). Голень и лапка обычно вооружены шипами. Голень, кроме того, может нести пару шпор (у Hesperiidae задняя голень с 2 парами шпор). На передней голени Раpilionidae и некоторых Hesperiidae имеется характерное лопастевидное образование - эпифиз (epiphysis), предположительно служащее для чистки усиков и глаз. Нормально развитая лапка состоит из 4-5 члеников. Дистальный членик вооружен парой коготков (ungues) и ложных коготков (paronychia) - удлиненных придатков, покрытых ресничками и волосками; между коготками расположена подушечка (pulvillus). Передние лапки у многих Papilionoidea подвержены значительной редукции, выражающейся в утрате сегментации, шипов, придатков дистального членика. Такая редукция характерна для самцов Danaidae, Satyridae, Nymphalidae и Lycaenidae, у которых передние ноги не функционируют при хождении. Передние лапки самок Satyridae, Danaidae и Nymphalidae, хотя и сохраняют сегментарное строение, не функционируют и всегда остаются прижатыми к груди (рис. 7).
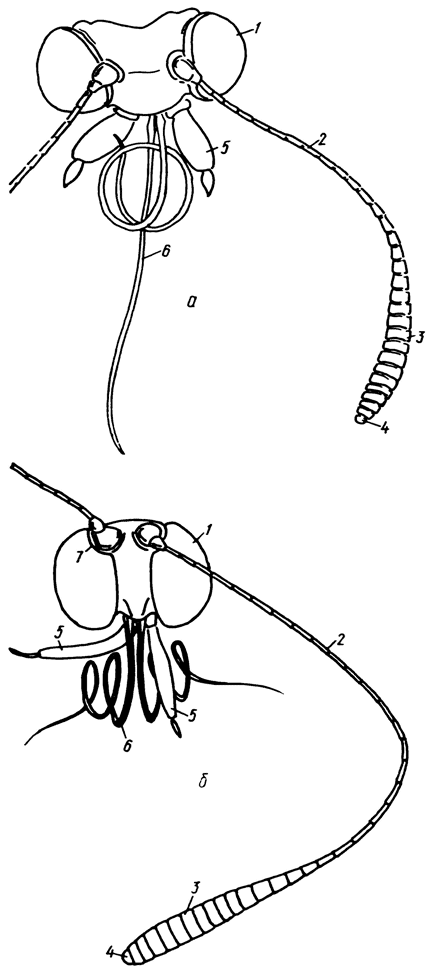
Рис. 5. Голова и ее придатки: а - Pyrgus malvae (L.) (Hesperiidae), б - Polyommatus icams (Rott.) (Lycaenidae) (1 - глаза, 2 - усики, 3 - булава усиков, 4 - вершина усиков, 5 - губные щупики, 6 - хоботок, 7 - вырезка глаза у основания усика)
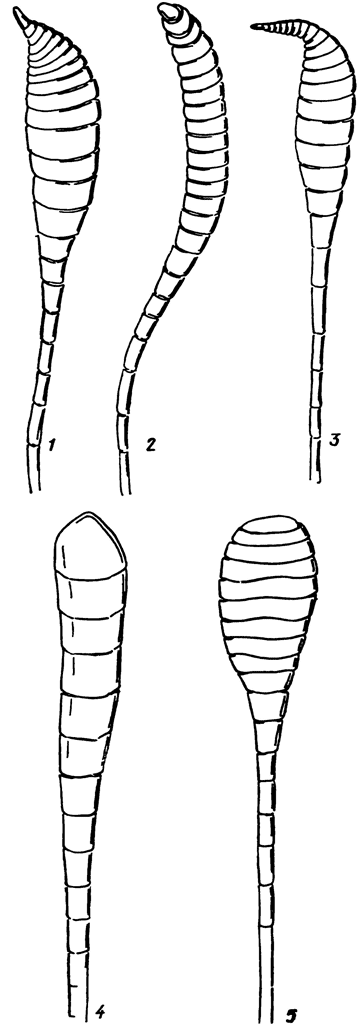
Рис. 6. Усики: 1, 2 - веретеновидные, булавовидные, 3 - крючковидные, 4 - 5 - головчатые

Рис. 7. Ноги: 1 - полное развитие всех трех пар (Papilio machaon L.), 2 - частичная редукция передней пары (Аrgynnis adippe (Rott.)
Крылья (alae) булавоусых чешуекрылых широкие, перепончатые, покрыты чешуями. Двухслойная перепонка, образующая крыло, поддерживается системой жестких жилок (venae), составляющих опорный аппарат крыла. По форме переднее крыло напоминает треугольник, заднее обычно округло-овальное. На крыле различают его основание, или корень (basis), вершину (apex) и задний, или анальный, угол (tornus), а также края: передний, или костальный (costa), внешний (termen) и задний, или анальный (dorsum). На заднем крыле эти образования выражены менее отчетливо, их пределы определяются по месту выхода к краю жилок радиального и анального комплексов (см. ниже). Внешний край крыла часто бывает волнистым или зубчатым, иногда он несет более или менее заметный вырост перепонки - хвостик. Описание рисунка привязывается к областям крыла: прикорневой, или базальной, дискальной, постдискальной, краевой, или маргинальной (субмаргинальное пространство находится в непосредственной близости от края, антемаргинальное - на некотором удалении от него), привершинной, или апикальной, и анальной; кроме того, различают верхнюю и нижнюю стороны крыла (рис. 8).
Система жилок крыла представлена у булавоусых чешуекрылых производными пяти продольных стволов (субкостального - Sc, радиального - R, медиального - М, кубитального - Сu и двух анальных - 2А и 3А), ветви которых выходят к костальному и внешнему краям. Основания медиальных ветвей соединены между собой и с ветвями R4+5 и Сu1 поперечными дискальными жилками D, замыкающими центральную ячейку крыла. У некоторых дневных бабочек дискальные жилки редуцированы, так что центральная ячейка оказывается незамкнутой. Применяемая в книге терминология жилкования (Miller, 1969) иллюстрируется рисунками в очерках семейств и сопровождается описанием характерных для родов особенностей. Жилкование переднего и заднего крыльев не совпадает, причем жилкование заднего крыла более постоянно и дает меньше диагностических признаков.
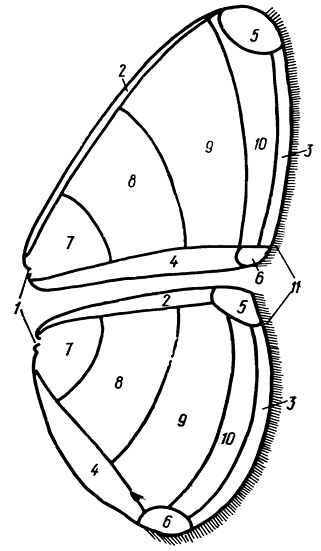
Рис. 8. Крылья: 1 - корень, 2 - костальный край, 3 - внешний край, 4 - анальный край, 5 - вершина, 6 - анальный угол, 7 - базальная (прикорневая) область, 8 - дискальная область, 9 - постдискальная область, 10 - маргинальная (краевая) область, 11 - бахромка
На переднем крыле неразветвленная субкостальная жилка Sc идет вдоль костального края и выходит к нему на большем или меньшем удалении от середины. Ствол R, ограничивающий спереди центральную ячейку, идет параллельно Sc, отдавая последовательно 5 ветвей - от R1 до R5; ствол R4+5 у всех Papilionoidea всегда разветвляется дистальнее центральной ячейки, у Неsperioidea все 5 ветвей R отходят непосредственно от центральной ячейки и не ветвятся. У некоторых Pieridae и всех Lycaenidae ветви R2 и R3 анастомозируют по всей длине, поэтому к краю крыла выходят не 5, а 4 ветви R. Локализация выхода к краю ветвей R3, R4 и R5 по отношению к вершине имеет важное диагностическое значение.
Медиальный ствол у всех дневных бабочек редуцирован, сохраняются лишь 3 ветви М1-М3. Кубитальный ствол, образующий заднюю границу центральной ячейки, дает 2 ветви Сu, и Сu2. Из 4 возможных в отряде анальных жилок А, признаваемых самостоятельными стволами, на переднем крыле Rhopalocera развита только 2А, самостоятельно начинающаяся у корня и идущая параллельно анальному краю к анальному углу. На заднем крыле субкостальная жилка Sc на всем протяжении слита с первой ветвью радиального ствола R1, образуя комплекс Sc + R1. Вблизи корня от Sc + R1, отходит короткий, иногда раздвоенный, рудимент плечевой, или прекостальной, жилки Н. Остальные ветви радиального ствола представлены здесь так называемым радиальным сектором Rs - неветвящейся жилкой, отходящей от медиального ствола М.
У всех дневных бабочек, кроме Раpilionidae, на заднем крыле имеется 2 анальные жилки 2А и 3А (у Papilionidae - только 2А).
Участки перепонки крыла, заключенные между жилками, называются ячейками. Ячейки обозначаются символами ограничивающих их жилок, соединенными тире (R5-М1, М1-М2 и т. д.). Пространство между радиальным и кубитальным стволами, замкнутое дискальными жилками, называется радиально-кубитальной, или центральной, ячейкой.
Обе поверхности крыльев покрыты окрашенными черепицеобразно налегающими друг на друга чешуями, образующими мозаичный рисунок. Различают 2 типа окраски чешуй: пигментный и структурный. В первом случае цвет чешуи определяется содержащимся в ней пигментом, во втором - оптической деятельностью модифицированной поверхности, на которой происходят явления интерференции. Структурные чешуи образуют элементы рисунка, имеющие металлический блеск. Иногда (у Gonepteryx) оптические структуры чешуй настолько тонки, что взаимодействуют не с видимой, а с ультрафиолетовой частью спектра, обусловливая возникновение так называемого скрытого рисунка. Чешуи, располагающиеся по внешнему краю крыльев, модифицированы и образуют бахромку (fimbrilla).
В крыловом рисунке различают фон - основную однородную окраску, занимающую всю или большую часть поверхности крыла, и расположенные по фону пятна, поля и перевязи. Пятном называется небольшой отличающийся от фона участок, расположенный в пределах одной ячейки. Пятно может быть полностью или частично окружено контрастной линией (оторочкой) и иметь в центре темную или светлую точку. Иногда пятно состоит из более или менее правильных концентрических колец, окружающих центральную точку или пятнышко,- в этом случае говорят о глазчатом пятне, или глазке. Мелкое пятно в зависимости от конфигурации называется точкой, штрихом или пестриной.
Поле представляет собой более обширный элемент рисунка, обычно имеющий размытые края, который располагается в пределах нескольких ячеек. Окраска полей может быть однородной и пестрой. Пятна часто образуют более или менее полные ряды (полый ряд возникает при наличии однородных пятен во всех ячейках крыла). При полном или частичном слиянии пятен одного ряда образуется перевязь - удлиненное поле, расположенное поперек жилок. Как и ряды пятен, перевязи могут быть полными, неполными и разорванными (при исчезновении одного или нескольких пятен в середине).
Положение полей, перевязей и пятен удобно привязывать к показанным на рис. 8 областям крыла. Положение пятен, кроме того, указывается по жилкам (дискальное пятно) и по ячейкам (пятна центральной ячейки, краевое пятно в ячейке Сu1 - Сu2 и др.). Используемая терминология рисунка обеспечивает опознавание видов региональной фауны, она предельно упрощена и, конечно, не может использоваться для филогенетических построений.
Брюшко удлиненное, у самцов конусовидное или уплощенное с боков, у самок веретеновидное, покрыто плотно прилегающими чешуями, иногда с рисунком. Из 10 сегментов, образующих этот отдел, у самцов нормально развиты 8, у самок 7. Каудальные сегменты редуцированы, а их придатки образуют наружные органы копулятивного аппарата (гениталии), признаки которых имеют первостепенное значение для диагностики и систематики. Нормальный сегмент брюшка образован дорсальным (тергит) и вентральным (стернит) полукольцами, соединенными в кольцо боковыми плейральными мембранами.
Гениталии самца (рис. 9) образованы модифицированными придатками IX и X брюшных сегментов и в состоянии покоя располагаются внутри VIII сегмента, тергит которого у некоторых видов также подвержен модификациям (жюльеновский орган Hipparchia и др.). IX брюшной сегмент представляет собой кольцо, в котором различаются расширенная куполовидная тергальная часть - тегумен (tegumen) и узкая стернальная часть - винкулум (vinculum). Вентральная часть винкулума часто несет более или менее обособленное булавовидное, пузыревидное или мешковидное образование - саккус (saccus). По бокам с тегуменом и винкулумом подвижно сочленяются двухслойные створки - вальвы (valvae), имеющие разнообразное, иногда сложное строение. Верхний, или костальный, и нижний, или вентральный, края вальвы обычно хорошо склеротизованы и по внутренней поверхности разделены мембранозным промежутком. Каудальный угол между краями вальвы называется ее вершиной. Часто мембранозный промежуток разделяет вершину вальвы на 2 отростка: костальный и каудальный. Внутренняя поверхность вальвы может нести один или несколько склеротизованных выростов разнообразной конфигурации - гарпы (harpae). У Hesperioidea вершинная часть вальвы бывает обособленной и носит название кукуллуса (cucullus).
Между вальвами, прикрепляясь к их базальной части, расположена юкста (juxta) - склеротизованный орган в виде пары более или менее удлиненных ветвей, поддерживающих эдеагус. X тергит в гениталиях самцов непарный, занимающий центральное положение или имеющий вид пары лопастей - ункус (uncus), у основания которого открывается трубка. X стернит представлен склеротизованным образованием - гнатосом (gnathos), чаще всего имеющим вид пары крючков или шиповатых пластинок - ветвей гнатоса (brachia). Лишь у некоторых Hesperiidae гнатос не имеет ветвей и является непарным образованием, расположенным под анальной трубкой. Наружная склеротизованная часть копулятивного органа - эдеагус (aedoeagus) - представляет собой трубку, иногда снабженную в проксимальной части, перед впадением семяизвергательного протока, слепым отростком - цекумом (coecum). Дистальная часть эдеагуса снабжена выворачивающейся перепончатой структурой - везикой (vesica), иногда несущей различной формы участки склеротизации - корнутусы (cornuti).
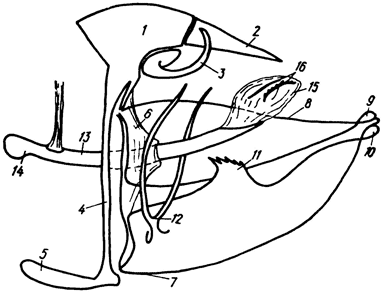
Рис. 9. Гениталии самца (схема): 1 - тегумен, 2 - ункус, 3 - ветви гнатоса, 4 - винкулум, 5 - саккус, 6 - диафрагма, 7 - вальва, 8 - костальный край вальвы, 9 - костальный отросток, 10 - каудальный отросток, 11 - гарна, 12 - юкста, 13 - адеагус, 14 - цекум, 15 - везика, 16 - корнутусы
Гениталии самки (рис. 10) образованы 3 последними сегментами брюшка (VIII-X). Самки булавоусых чешуекрылых имеют 2 половых отверстия: копулятивное, расположенное между VII и VIII стернитами, и яйцевыводное - на конце брюшка. Копулятивное отверстие (ostium bursae) располагается на интерсегментальной мембране в окружении более или менее обособленных участков склеротизации, которые в соответствии с их положением называются антевагинальной и поствагинальной пластинками (lamellae ante- et postvaginalis). Строение их и топографические взаимоотношения у представителей различных семейств и родов отличаются значительной сложностью, вместе с тем характер склеротизации этой области часто является ключевым диагностическим признаком.
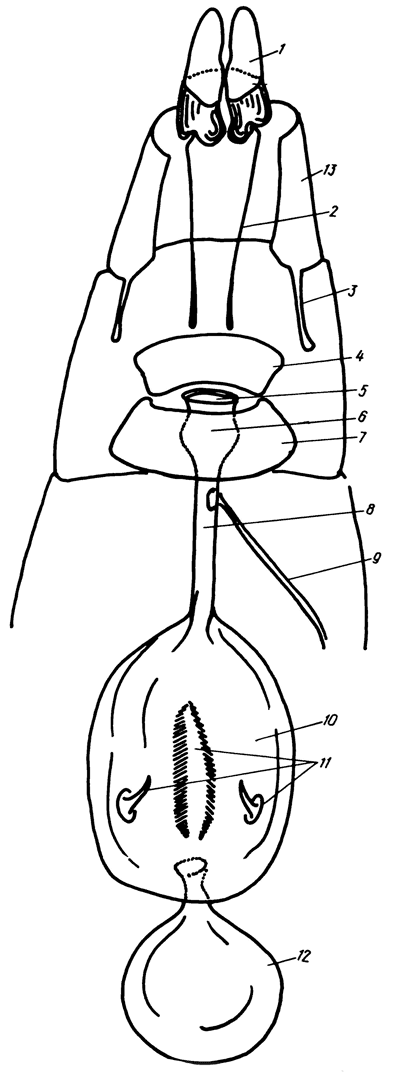
Рис.10.Гениталии самки (схема): 1 - анальные сосочки, 2 - задние апофизы, 3 - передние апофизы, 4 - поствагинальная пластинка, 5 - остиум, 6 - антрум, 7 - антевагинальнал пластинка, 8 - дуктус, 9 - семенной проток, 10 - бурса, 11 - сигнумы, 12 - аппендикс бурсы, 13 - VIII тергит
При копуляции половые продукты поступают в антрум (antrum) - расширенную, иногда склеротизованную воронку, соединенную с протоком копулятивной сумки - дуктусом (ductus bursae copulatrix) - перепончатой или более или менее полно склеротизованной трубкой. Дуктус впадает в мешкообразную перепончатую совокупительную сумку (bursa copulatrix), стенка ее может быть гладкой или складчатой, иногда она инкрустирована различной формы склеритами - сигнумами (signа). У Pieridae бурса снабжена стебельчатым аппендиксом (appendix bursae). От дуктуса берет начало семенной проток (ductus seminalis), ведущий к яйцевыводному протоку. Устье семенного протока на дуктусе иногда располагается в центре склеротизованной бляшки.
Яйцевыводная часть гениталий самок булавоусых чешуекрылых имеет более однообразное строение. Здесь различают модифицированный VIII тергит, от переднего края которого внутрь полости тела отходят парные, направленные орально, передние апофизы (apophyses anteriores), а также анальные сосочки (papillae anales) - парные полупрозрачные покрытые щетинками лопасти, расположенные на X сегменте, между которыми открываются яйцевыводное и анальное отверстия. IX сегмент просматривается в виде охватывающей сосочки проксимальной, или базальвой, зоны склеротизации, от которой берут начало задние апофизы (apophyses posteriores).
Андрокониальные структуры представляют собой группы специализированных чешуй - андроконий (androconia), которые служат для испарения пахучего секрета гиподермальных желез у самцов. У дневных бабочек андрокониальные чешуи чаще всего локализуются на верхней стороне передних крыльев (у Danaidae - на задних крыльях), располагаясь между покровными чешуями или образуя различных размеров и формы андрокониальные поля, выделяющиеся на фоне крыла цветом и фактурой (мучнистые, бархатистые, атласные). У некоторых Hesperiidae на переднем крыле развит так называемый костальный заворот - завернутая на верхнюю сторону лопасть перепонки крыла, под которой располагается кисть удлиненных андроконий. Кроме крыльев, андрокониальные структуры у дневных бабочек могут располагаться на брюшке (характерные выворачивающиеся кисти Danaidae; вероятно, что эту же функцию выполняют модифицированные чешуи жюльеновского органа Hipparchia), а также на голенях задних ног (тибиальная кисть некоторых Hesperiidae). Признаки андрокониального аппарата - форма и размеры андрокониальных чешуй, локализация и характер андрокониальных полей, а в некоторых случаях и воспринимаемый человеком аромат - в отдельных группах бабочек имеют важное диагностическое значение (Кузнецов, 1915, 1929; Hering, 1926; Яхонтов, 1935, 1939; Ehrlich, 1958; Klots, 1970; Higgins, 1975; Фалькович, Стекольников, 1978).
Таблица для определения семейств по внешним признакам
1(2). Голова широкая, ее поперечный размер явно больше продольного, глаза
широко раздвинуты, между ними имеется хохолок из волосовидных чешуй, жилки R переднего крыла отходят непосредственно от центральной ячейки, не ветвятся ...................Hesperiidae (ч. II)
2(1). Голова округлая, ее поперечный размер примерно равен продольному, глаза занимают почти всю поверхность головной капсулы, сдвинуты близко, некоторые жилки R переднего крыла ветвятся перед выходом к краю.
3(6). Все ноги у обоих полов нормально развиты и функционируют при хождении.
4(5). Заднее крыло с одной жилкой А ...........Papilionidae (с. 47)
5(4). Заднее крыло с двумя жилками А ............Pieridae (с. 64)
6(3). Передние ноги у самцов или у обоих полов развиты не полностью и не функционируют при хождении.
7(10). Передние ноги редуцированы только у самцов.
8(9). Глаза окаймлены светлыми чешуйками, имеют вырезку у основания усиков, губные щупики короче груди ..........Lycaenidae (ч. II.)
9(8). Глаза без светлого окаймления, без вырезки у основания усиков, губные щупики равны длине груди ............Libytheidae (ч. II)
10(7). Передние ноги редуцированы у обоих полов.
11(12). Андрокониальное поле расположено на заднем крыле ...................Danaidae (с. 187)
12(11). Андрокониальное поле расположено на переднем крыле или отсутствует.
13(14). Одна или несколько жилок у корня переднего крыла вздуты ..............Satyridae (с. 105)
14(13). Вздутых жилок на переднем крыле нет .....Nymphalidae (ч. II)
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://butterflylib.ru/ 'Чешуекрылые, бабочки, мотыльки, моли'