Colias palaeno L. - Торфяная желтушка
Распространена в Европе, в Альпах и по северу Евразии доходит до Дальнего Востока, населяя Якутию, Магаданскую обл., Камчатку и о. Сахалин. В Приамурье, Приморье и в Забайкалье встречается спорадически, придерживаясь заболоченных участков и лиственничных марей в долинах рек. Еще южнее ее популяции становятся обитателями высокогорных тундр альпийской зоны. Спорадически, придерживаясь лишь наиболее высоких одиночных вершин или групп вершин, эта желтушка известна теперь, но нашим исследованиям, в гольцах южного Сихотэ-Алиня, а по литературным данным, известна из альпийской зоны больших островов Японии и высоких горных массивов Чаньбошаня в северной части п-ова Корея (Esaki, 1929; Seok, 1934, 1941; Esaki a. Yokoyama, 1955). На крайнем северо-востоке Сибири доходит до северной границы Чукотско-Анадырской тундры. Отсюда она распространена и дальше к востоку - в зоне тундры и лесотундры Северной Америки.
Естественно, занимая столь обширный ареал с различными эколого-географическими условиями, этот вид в морфологическом отношении не может оставаться монотипным. При обработке имеющегося в нашем распоряжении довольно большого материала по этому виду, собранного как мною лично в различных частях Дальнего Востока, так и полученного от отечественных и заграничных коллег-лепидоптерологов, я пришел к выводу, что изучение внутривидовой систематики у С. palaeno с учетом экологии и распространения ее таксонов представляет большой теоретический интерес.
Я считаю, что на Дальнем Востоке встречается не менее шести подвидов торфяной желтушки. На крайнем северо-востоке Сибири распространен подвид С. р. schippewa Edw. (рис. 25), который раньше указывался для Аляски и северо-западных арктических частей Канады (Freeman, 1958а; Legge, 1965). По сравнению с другими дальневосточными географическими расами этого вида он отличается наименьшими размерами (35-36 мм) и неширокой краевой каймой. У самок, окрашенных в палево-белый цвет, бурая краевая кайма очень широкая у апекса передних крыльев и едва сохранившаяся или узкая у верхнего края задних крыльев. Некоторые американские авторы (Holland, 1949) приводят С. р. schippewa как самостоятельный вид наряду с распространенным также на Аляске видом С. palaeno. Нам представляется более правильным, как это и высказывалось другим американским энтомологом (Klots, 1958), что в арктической Америке от Аляски до р. Мекензи и Большого Невольничьего озера встречается только С. р. schippewa. Однако считать ли его подвидом или согласиться с мнением В. Голланда, выделившего С. р. schippewa в самостоятельный вид.
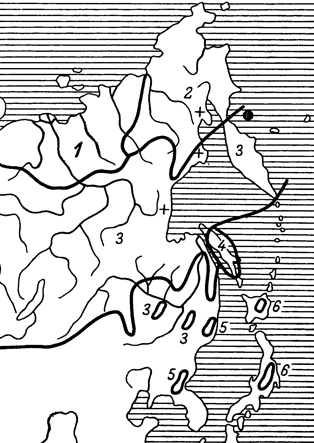
Рис. 25. Ареалы дальневосточных подвидов Colias palaeno. 1 - С. р. arctica; 2 - C. р. schippewa; 3 - С. р orientalis; 4 - С. р. sachalinensis; 5 - С. р. sugitani; 6 - С. р. aias. Крестики - сохранившиеся редкие местонахождения подвида С. р. arctica вне его основного ареала; черный кружок - единственное нахождение на о. Карагинском популяции С. р. arctica
Полученные нами новые данные по систематике, экологии и распространению этой бабочки на сибирском отрезке ее ареала вносят, как нам кажется, известную ясность в решение этого вопроса. По окраске сибирские экземпляры ничем существенным не отличаются от североамериканских. Гениталии их самцов по сравнению с другими сибирскими подвидами (рис. 26) имеют то отличие, что у сибирских С. р. schippewa псеудоункус очень укорочен, как и у аляскинских особей (Petersen, 1963). Принимая во внимание, как показал этот исследователь, адаптивное значение названной детали в строении гениталий, а следовательно, и важность ее для систематики, нам кажется, что эти признаки у С. р. schippewa ставят его в морфологическом отношении выше, чем категория подвида. Таксономически мы отнесли бы его к систематической единице полувид (Завадский, 1967), но еще правильнее применить к нему название vicespecies, предложенное еще раньше А. Авиловым (1913) для подобных же таксономических понятий у некоторых видов рода Parnassius. Этот термин выражает точнее состояние почти завершающегося процесса видообразования, что и подтверждается эколого-географическими особенностями этой бабочки. Ее ареал охватывает Чукотский полуостров до устья р. Колымы. На западе он граничит с ареалом подвида С. р. arctica Vrty., а на юге - с ареалом подвида С. р. orientalis Stg. Переходные экземпляры очень редки и отмечены только на юге ареала, где, как это наблюдалось нами, идет в настоящее время процесс активного проникновения С. р. chippewa в ареал С. р. orientalis. Если последний подвид является только обитателем долинных моховых болот и лиственничных редкостоев, то подвидом С. р. schrippewa осваиваются различные биотопы от долинных марей и до каменистых тундр высокогорных ландшафтов. Вероятно, тот же процесс внедрения С. р. schippewa в ареал подвида С. р. artica происходит и у западных границ его распространения. Во всяком случае в Заполярье (Анюйский хребет), восточнее низовьев р. Колымы, мы уже не встретили ни разу С. arctica, тогда как С. р. schippewa была обычной бабочкой в долинных и горных тундрах. Доказательством смещения ареала С. р. arctica к западу под воздействием прогрессирующего распространения С. р. schippewa являются редкие находки арктического подвида на ареале как С. р. schippewa, так и на ареале С. р. orientalis близ Охотского моря и на о. Ольском у берегов последнего.
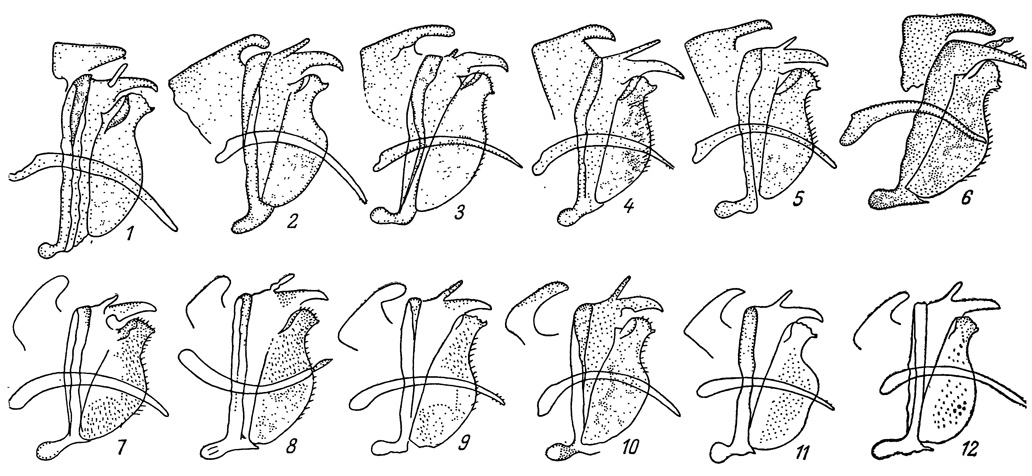
Рис. 26. Гениталии самцов дальневосточных подвидов Colias palaeno. 1 - С. р. arctica, о. Карагинский; 2 - С. р. orientalis, Магаданская обл.; 3 - С. р. schippewa, Магаданская обл.; 4 - С. р. orientalis, Забайкалье; 5 - С. р. aias, альпийская зона о. Хонсю; 6 - С. р. sugitani, альпийская зона п-ова Корея; 7 - С. р. schippewa, сев.-зап. Канада; 8 - С. р. arctica, Нижнеколымск; 8 - С. р. schippewa, Анюйский хребет; 10 - С. р. orientalis, северная Монголия; 11 - С. р. sachalinensis, о. Сахалин; 12 - с. р. arctica, о. Ольский (Магаданская обл.)
В связи со сказанным о продолжающемся процессе расселения С. р. schippewa необходимо еще отметить нахождение несколько своеобразной формы С. р. palaeno на Карагинском острове у северо-восточных берегов Камчатки. По величине она (40 мм) приближается к С. р. arctica. Общая оранжевая окраска самцов менее яркая, чем у С. р. arctica и С. р. schippewa. Краевая черная полоса у них уже, особенно на задних крыльях, и имеет с внутренней стороны на передних крыльях тупые зубчики. Весь низ задних крыльев густо покрыт темными чешуйками. Кроме того, у самцов заметно заострены у апекса передние крылья (рис. 27). Самки почти ничем не отличаются от самок С. р. arctica.

Рис. 27. Дальневосточные подвиды Colias palaeno. 1 - С. р. orientalis; 2 - С. р. arctica; 3 - С. р. arctica f. haraganica; 4 - С. р. aias; 5 - С. р. schippewa; 6 - С. р. sugitani. Верхний ряд - самцы; нижний - самки
Я имею всего три бабочки этой формы (два самца и самка), собранные 14 июля, 4 и 30 августа в 1929 г. В. И. Разумовским. Вероятно, эта форма является измененной островными условиями популяцией подвида С. р. arctica, ранее более широко распространенного в северо-восточной Сибири, но позднее вытесненного С. р. sphippewa. Эта популяция заслуживает выделения ее в особую форму арктического подвида С. р. arctica Vrty. f. karaganica nova.
Периоды лёта у С. р. arctica и С. р. schippewa совпадают и происходят со второй декады июля и до конца августа.
Южнее С. р. arctica и С. р. schippewa особенно широким распространением пользуется подвид С. р. orientalis, встречаясь от Забайкалья через южные районы Якутии и Магаданской обл. до Камчатки включительно. Обычен и в бассейне Амура, но при наличии характерных стаций - лиственничных редкостойных марей с моховым покровом и зарослями голубики (Vaccinium uliginosum) - его кормового растения.
По размерам бабочки определенно крупнее подвидов С. р. schippewa и С. р. arctica, они лимонно-желтые, имеют и более широкую краевую полосу. Самки зеленовато-белые, снизу на задних крыльях густое опыление бурых чешуек. У самцов общий фон снизу крыльев желтый с густым покровом зеленых чешуек на задних крыльях.
К югу от Амура этот подвид становится реже, но по горным системам Большого Хингана, Восточно-Маньчжурской горной страны и Сихотэ-Алиня все еще проникает до 46-47° с. ш., оставаясь всегда верным спутником долинных и горных моховых марей. В пределах своего обширного ареала этот подвид не остается неизменным и образует несколько отличающихся популяций. Так явно выделяются забайкальские экземпляры с очень длинным псеудоункусом у самцов (рис. 26); монгольские особи меньше забайкальских, у самок краевая черная полоса у апекса имеет светлые штрихи; самцы из среднего Сихотэ-Алиня отличаются более яркой окраской и несколько вытянутыми вдоль передними крыльями.
Период лёта у этого подвида продолжается со средины июня и до второй половины июля. Гусеницы появляются в августе на голубике (Vaccinium uliginosum).
Южнее, уже в высокогорных условиях южного Сихотэ-Алиня и северных частей п-ова Корея, изолированно обитает подвид С. р. sugitani Esaki, а в альпийском поясе гор Центральной Японии - подвид С. р. aias Fruhst. Наконец, на о. Сахалин популяция торфяной желтушки выделена (Matsumura, 1925) также в подвид С. р. sachalinensis Mats.
По сравнению с С. р. orientalis каждый из этих подвидов в той или другой степени отличается от него. Резкое отличие имеет С. р. aias, географически наиболее изолированный подвид. По размерам (50 мм) он является самым крупным подвидом этой желтушки. Самец ярко-желтый, самка зеленовато-белая. Краевая полоса у обоих полов очень широкая, занимает третью часть поверхности крыльев (рис. 27). Вероятно, этот подвид давно изолирован от всех других популяций вида.
С. р. sugitani также довольно крупный подвид (47-48 мм), занимает среднее положение между подвидами С. р. orientalis и С. р. aias. Окраска самцов лимонно-желтая, но бледнее, чем у подвида С. р. orientalis, а краевая черная кайма почти вдвое шире, чем у последнего. У зеленовато-белых самок на черной кайме имеется несколько продольных светлых штрихов. Экологически С. р. aias и C. p. orientalis представляют большой интерес. Их можно считать характерными представителями альпийской фауны. Ряд японских энтомологов (Esaki, 1929; Uchida, 1929, 1930) приводят эти подвиды для высоких вершин о. Хонсю. Полученные мною от К. Тамануки несколько экземпляров С. р. aias, собранных на горе Эбоши 21 июля 1931 г. этого же острова, оказались очень характерными для этого подвида. С них и сделан рисунок (рис. 27). В горах п-ова Корея в прежние годы (Seok, 1934, 1941) подвид С. р. sugitani найден и описан с вершин Пайктусана и горного массива Камбо, подымающихся до 2000 м над ур. м. и выше. По моей просьбе на горе Пайктусан корейский энтомолог Дю Дон Юр собрал в начале августа 1960 г. несколько экземпляров этого подвида. Мною этот подвид был собран только однажды 15 июля 1949 г. на вершине горы Цамо-Дынза (1750 м над ур. м.) в южном Сихотэ-Алине в условиях каменистых тундр.
Сахалинский подвид - С. р. sachalinensis вопреки мнению авторов сводки о дневных бабочках острова (Hori et Tamanuki, 1937), считавших название этого подвида синонимом С. р. orientalis, все же заслуживает своего выделения. Указания этих авторов на то, что С. р. sachalinensis очень изменчив и дает якобы переходы к С. р. orientalis, не всегда могут быть приняты во внимание уже по одному тому, что в зонах интерградации между двумя близкими подвидами всегда встречаются и промежуточные между ними варианты. Во всяком случае среди сахалинской популяции торфяной желтушки доминируют самцы, имеющие палевожелтую окраску сверху крыльев и сильно опыленный бурыми чешуйками низ задних крыльев. Вот за ними и необходимо сохранить предложенное название С. р. sachalinensis (Matsumura, 1925). Кроме того, при выделении географических и экологических подвидов необходимо принимать во внимание характер изолирующих популяцию факторов и ее исторический возраст. Про сахалинский подвид торфяной желтушки можно сказать, что он, несомненно, молодой подвид, генетически еще не потерявший связи с исходным подвидом С. р. orientalis, несмотря даже на свое островное обособление.
Из сказанного о дальневосточных подвидах С. р. palaeno можно сделать следующее заключение. Внутривидовая изменчивость у различных форм С. р. palaeno в связи с их экологией и распространением дает богатый материал для изучения вопросов микроэволюции у бабочек. На приведенных примерах распространения подвидов С. palaeno нетрудно было уловить ту закономерность изменчивости, что и у подвидов, имеющих северные ареалы: уменьшается ширина каймы сверху на крыльях, тогда как при распространении подвидов к югу, наоборот, возрастает ширина краевой полосы, достигая у С. р. alas наибольшего выражения.
Исторически географические подвиды у С. palaeno возникли в различное время. Поэтому у одних подвидов (например, у С. р. schippewa и С. р. alas) с явно выраженной географической изоляцией и резко обособленной морфологической структурой мы наблюдаем почти законченный процесс становления нового вида. Поэтому по происхождению мы можем считать их более древними. У других подвидов, возникших в более позднее время, эколого-географическая их изоляция еще не закончилась полностью, а морфологическое строение оказывается недостаточно стойким.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://butterflylib.ru/ 'Чешуекрылые, бабочки, мотыльки, моли'