Географическое распространение чешуекрылых Кашка-Кума
В вахшских песках зарегистрировано 119 видов чешуекрылых. Общий список высших чешуекрылых песчаного массива и распространение видов в трех участках песков и других основных биотопах представлен в табл. 6. Однако в числе отмеченных некоторые виды оказались в песчаной пустыне явно случайно, залетев из близ расположенных тугаев или возделываемых земель, и они в песках не обитают. Таких залетных видов 29:

Если исключить из общего списка (табл. 6), эти посторонние для песков элементы, помеченные в таблице звездочкой (*), то остается 90 видов высших чешуекрылых, или 30% от общего их числа в долине. В дальнейшем анализе эти залетные виды не учитываются, и речь будет идти только о формах, размножающихся в песках (т. е. о 90 видах).
Некоторые поливольтинные виды в песках встречаются лишь в определенные сезоны, и в благоприятное время года (весной) могут нормально развиваться на травянистой растительности, а в дальнейшем мигрируют на ближайшие участки тугайных зарослей, поливные земли или, быть может, даже в горы, где и происходит развитие остальных генераций. К числу таких форм относятся Pieris rapae L., Colias erate Esp. и Pyrameis cardui L., характерные в долине для культурного ландшафта и тугаев. Возможно, к этой группе принадлежит и Satyrus enervata Alph., по-видимому, откочевывающая летом в горы, где и летует в укромных местах. Ряд видов (Hypermnestra helios Nick., Apharitis epargyros Ev., Neoris stoliczkana schencki, Chilena sordida Ersch., Cucullia argentina F., Pericyma albidentaria Frr., Melicleptria scutosa Schiff., Rhynchodontodes ravalis H. S., Microloxia herbaria Ld. и некоторые другие) развивается на растениях, не характерных для вахшских песков, растущих небольшими скоплениями, главным образом, на глинистых обнажениях, в понижениях на уплотненной почве, на окраине песков. К числу подобных случайных для Кашка-Кума растений принадлежат верблюжья колючка (Alhagi canescens), виды полыни (Artemisia), ZygophyllumGontscharovii и некоторые другие. В низовьях Кафирнигана и Сурхан-Дарьи из таких растений в пески вторгается также и степная мимоза Prosopis stephaniana со специфичными для нее видами бабочек. Но как бы то ни было, чешуекрылые, связанные с этими растениями, обитают в песках, хотя совсем не характерны для них.
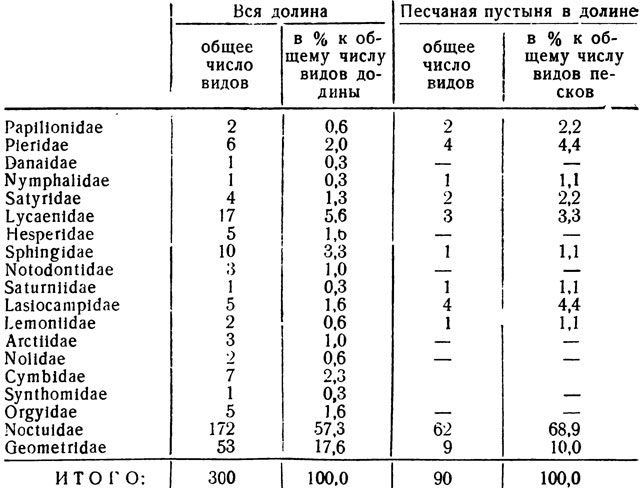
В Вахшской долине вообще бедно представлены многие группы чешуекрылых: все семейства дневных бабочек - Rhopalocera (особенно Nymphalidae), Notodontidae, Arctiidae, Orgyidae. Фауна же бабочек песков в сравнении с долинной в целом или другими ее биотопами оказывается еще более обедненной. Почвенные и жесткие климатические условия, главным образом, режим влажности оказываются непреодолимым барьером для проникновения в пески мезофитной растительности. Поэтому все без исключений монофаги и олигофаги, связанные с мезофильными и тем более с гигрофильными растениями, в песках отсутствуют. Из фауны Вахшских песков совершенно выпадают представители ряда семейств (Danaidae, Arctiidae, Synthomidae, Nolidae, Cymbidae и Orgyidae), хотя вообще-то в долине некоторые виды из этих групп имеются. Характерно также, что в пески не проникают не только северные формы, но полностью отсутствуют и тропические элементы, встречающиеся в других биотопах долины. В табл. 1 показано численное распределение видов по семействам как в песках, так и в долине вообще. Из приведенных цифр видно, что фауна песков отличается поразительно бедным видовым составом большинства семейств (при этом ряд групп, как сказано, вообще здесь не представлен); с другой стороны, обращает на себя внимание относительное обилие здесь совок - сем. Noctuidae, число которых (62 вида) составляет около 70% (68, 9%) всего видового состава высших чешуекрылых песков. В то же время Noctuidae всей Вахшской долины по отношению к общему числу ее высших бабочек (300 видов) составляют относительно значительно меньшую величину- 57,3% (172 вида). Из мелких групп высших бабочек в песках представлены сравнительно хорошо шелкопряды - Lasiocam pidae (4/5 от общего числа видов в долине) и белянки - Pieridae (4 вида из 6). Пяденицы (Geometridae) по числу видов среди высших чешуекрылых в долине и в песках занимают второе место после Noctuidae; однако 9 видов - это лишь 10% от общего числа песчаных форм, тогда как по долине в целом пяденицы составляют 17,6% (53 вида).
Такая исключительная обедненность пустынной фауны объясняется тем, что большинство среднеазиатских видов Rhopalocera и многих семейств Heterocera является в той или иной степени мезофилами, или они, не будучи сами таковыми, очень тесно связаны с мезофитной растительностью (монофаги, олигофаги). Примечательно, что из обширного семейства Nymphalidae не только в песках, но и во всей Вахшской долине встречается лишь одна космополитическая Pyrameis cardui L. Нет ни одного представителя родов Vanessa, Melitaea, Argynnis. Характерно также, что такие громадные группы, как Geometridae и (из низших) Tortricidae, виды которых связаны преимущественно с древесной мезофильной растительностью, р. песках (да и во всей долине) представлены ничтожно малым числом форм. Только Noctuidae и из группы низших бабочек огневки - Pyralidae, содержащие множество ксерофильных форм и ведущие в фазе гусеницы, как правило, скрытный образ жизни, численно преобладают как вообще в пустынных местностях Средней Азии, так и в особенности в песках. В этих группах очень многие формы не только физиологически приспособились к обитанию в крайне неблагоприятных условиях пустынной зоны, но у них в течение эволюционного развития соответственно изменился образ жизни и появились некоторые адаптивные морфологические структуры. В семействе Noctuidae наиболее многочисленными и характерными для песков являются виды трех родов: Anumeta, Armada и Leucanitis; из Pyralidae в песках особенно хорошо представлено подсемейство Phycitinae, в частности, большой род Heterographis, характерный для пустынной зоны.
В современном географическом распространении видов в известной степени отражаются как их история, происхождение, так и экологические потребности в настоящее время. Однако, какой из этих факторов является главным в формировании современного ареала вида, исторический или экологический? В этом вопросе нет единства представлений ни у зоологов, ни у ботаников. Некоторые ботаники и зоологи считают что именно история флоры и фауны является решающим моментом в формировании ареалов, а экологический фактор играет второстепенную роль и определяет лишь распространение форм по биотопам внутри исторически сложившегося ареала (как будто можно рассматривать историю видов вне экологических факторов!). Однако подобные вопросы могут быть разрешены не путем создания теоретических предпосылок, из которых исходят при рассмотрении материала, а наоборот, путем первоначального анализа фактов, который в дальнейшем может послужить для теоретических обобщений. Ввиду того, что работ по зоогеографии чешуекрылых очень мало, (по Средней Азии вообще нет), а принципы зоогеографического районирования горных стран в лепидоптерологии не разработаны, то при написании данного и последующих двух разделов на материале пустынной фауны будет рассмотрен и этот, последний, вопрос, без правильного решения которого невозможно выяснить истинное распределение организмов в области со столь сложными и пестрыми горными сооружениями, какими характеризуется Средняя Азия.
Таблица 1
Для выяснения распространения кашкакумских бабочек необходимо было объединить виды в ряд групп то типам ареалов и выяснить степень зависимости распространения видов от экологических условий и связи фауны Средней Азии с другими областями Палеарктики. При этом важно было рассмотреть и вертикальное распределение чешуекрылых.
Как не велико разнообразие ареалов даже среди форм лишь одного биотопа, в данном случае песков, оказалось возможным свести их к 12 основным типам, или группам видов:
I. Эндемики Средней Азии.
II. Распространенные от западных районов Средней Азии до Монголии или восточных районов Джунгарии.
III. Общие для Средней Азии, Кашгарии и Индии.
IV. Общие для Средней Азии, Афганистана и Белуджистана.
V. Общие для Средней Азии Афганистана, Ирана и Ирака.
VI. Общие для Средней Азии, Афганистана, Ирана, Передней и Малой Азии, Закавказья или также юго-востока Европейской части СССР.
VII. Общие для территории, простирающейся от Средней Азии и Афганистана до восточного побережья Средиземного моря и Закавказья, а также до Северной Африки.
VIII. Общие для Средней Азии и Закавказья.
IX. Общие для Средней Азии, юго-востока Европейской части СССР или также для Южного Приуралья и северовосточной части Кавказа.
X. Общие для территории, простирающейся от Средней Азии и Афганистана до Южной Европы, Передней и Малой Азии или также до Северной Африки.
XI. Распространенные в Палеарктике очень широко.
XII. Голаркты и космополиты.
Рассмотрим каждую из этих групп в отдельности.
I ГРУППА. ЭНДЕМИКИ СРЕДНЕЙ АЗИИ
В этот господствующий по числу видов комплекс входит 32 вида, или 35,6% от общего количества высших чешуекрылых фауны песков. Сюда вошли и виды, ареал которых захватывает также Кульджинский край в Северо-Западном Китае и северную пограничную полосу Афганистана или Северо-Восточного Ирана, поскольку в фаунистическом отношении эти области являются составными частями Средней Азии. Однако, все перечисленные формы не идут севернее южной окраины Усть-Урта и южного побережья Аральского моря.
Подавляющее большинство эндемиков (24 вида) распространено только в западной части Средней Азии* и не встречается восточнее Ферганской долины. В списке они отмечены звездочками (*). Из них Thargelia arenicola Stshetk. и Cidaria tadzhikaria Stshetk. пока известны как очень локальные виды.

* (Здесь и в дальнейшем под западной частью Средней Азии подразумеваются ее районы к западу от восточной окраины Ферганской долины.)
Большинство рассматриваемых форм распространено в равнинной полосе и низинных долинах Средней Азии. Некоторые виды поднимаются в предгорья или низкогорья до 700 - 800 м над ур. м. и только Paraegle tesselata местами доходит до 2000 м. Почти 2/3 видов (20) являются типичными теплолюбивыми псаммофилами. Остальные также приурочены к пустынным биотопам, в которых видное место занимает или даже господствует эфемеровая растительность. Все формы, кроме Porphyrinia gratiosa, моновольтинные.
II ГРУППА. ВИДЫ, АРЕАЛЫ КОТОРЫХ ОХВАТЫВАЮТ БОЛЬШУЮ ЧАСТЬ СРЕДНЕЙ АЗИИ, А ТАКЖЕ СЕВЕРО-ЗАПАДНУЮ МОНГОЛИЮ ИЛИ ДЖУНГАРИЮ, - 2 ВИДА (2,2%):
Scythocentropus scripturosa
Diadochia malitiosa (ssp. usurialis)
Равнинные пустынные виды, из которых второй является псаммофилом; подвид usurialis Pglr. эндемичен для западной части Средней Азии. Возможно к этой категории принадлежит и ряд других, особенно псаммофильных форм, но песчаная фауна Монголии до сих пор остается почти неизвестной. Виды этого комплекса по распространению и экологической характеристике близки группе эндемиков Средней Азии.
III ГРУППА. ВИДЫ, ОБЩИЕ ДЛЯ СРЕДНЕЙ АЗИИ, ИНДИИ, КАШГАРИИ, СЕВЕРНЫХ РАЙОНОВ АФГАНИСТАНА И ИРАНА. - 1 вид (1,1%):
Neoris stoliczkana (ssp. schencki)
Большая часть ареала N. stoliczkana приходится на горную Среднюю Азию до Юлдуса и Зайсана включительно; генетически этот вид явно тяготеет к ориентальной фауне; встречается он кое-где в Вахшских песках, но характерен для горных районов. Моновольтинный полифаг.
IV ГРУППА. ФОРМЫ, ОБЩИЕ ДЛЯ СРЕДНЕЙ АЗИИ, АФГАНИСТАНА И БЕЛУДЖИСТАНА-8 ВИДОВ (8,9%):

Четыре вида, помеченные звездочкой (*), в Средней Азии встречаются только в западной ее части. Ареал Epinephele narica в основном очерчен указанной территорией, но на северо-востоке захватывает также пустынные районы южной части левобережья р. Урал; однако его подвид fusca Stshetk. эндемичен для юго-западной части Средней Азии. Ареалы Neolycaena tengstroemi и Satyrus enervata на востоке выходят за пределы Средней Азии и простираются до Западной Монголии. Оба вида - равнинно-горные ксерофиллы; в горах поднимаются до 3000 м и встречаются, главным образом, на сухих склонах. Остальные представители группы являются типичными обитателями пустынных биотопов равнин и низинных долин, некоторые заходят в предгорья или пустынные низкогорья, но не выше 900 м над ур. м. Из числа низинных форм Leucanitis sinuosa и Atomorpha hedemanni встречаются только в песках.
V ГРУППА. ВИДЫ, ОБЩИЕ ДЛЯ СРЕДНЕЙ АЗИИ, АФГАНИСТАНА, ИРАНА И ИРАКА-5 ВИДОВ (5,6%):

Два вида этого комплекса в своем распространении не выходят на востоке за пределы западной части Средней Азии (Margelana versicolor, Sidemia apotheina), ареалы остальных простираются до восточных пределов Средней Азии, причем Apharitis epargyros идет еще дальше, до Монголии. Одни формы характерны для жарких низинных долин и равнин (Apharitis epargyros, Armada limata), Armada hueberi обыкновенна также в пустынных предгорьях и низкогорьях, а два остальных довольно многочисленны от равнинных пустынь до 2500 м над ур. м., хотя и в горах предпочитает ксерофильные биотопы.
Рассматриваемая группа предсгавлена одним типичным псаммофилом (Armada limata), тремя эромофилами и одним равнинно-горным ксерофилом (Margelana versicolor).
VI ГРУППА. ВИДЫ, ОБЩИЕ ДЛЯ СРЕДНЕЙ АЗИИ, АФГАНИСТАНА, ИРАНА, ПЕРЕДНЕЙ И МАЛОЙ АЗИИ, ЗАКАВКАЗЬЯ ИЛИ ТАКЖЕ ДЛЯ ЮГО-ВОСТОКА ЕВРОПЕЙСКОЙ ЧАСТИ СССР - 5 ВИДОВ (5,6%):

Chondrostega pastrana и Paraegle ochracea характерны для эфемеровой растительности пустынных подгорных равнин, нижней полосы низкогорий и закрепленных песков. В Средней Азии они встречаются только в ее западной части и не доходят до юго-востока Европейской части СССР Остальные три вида распространены значительно шире: на северо-запад до юго-востока Европейской части СССР, в Средней Азии - до ее восточных окраин или до Западной Монголии (Leucanitis flexuosa). При этом по вертикали по наиболее сухим склонам L. flexuosa доходит до 800 м, Lasiocampa eversmanni и Cucullia boryphora поднимаются довольно высоко в горы (соответственно до 2900 и 2300 м). В рассматриваемой группе большинство составляют пустынные виды, т. е. эремофилы (но нет псаммофилов). Только Lasiocampa eversmanni принадлежит к равнинно-горным ксерофилам.
VII ГРУППА. ВИДЫ, АРЕАЛ КОТОРЫХ ОХВАТЫВАЕТ ТЕРРИТОРИЮ От СРЕДНЕЙ АЗИИ И АФГАНИСТАНА ДО ВОСТОЧНОГО ПОБЕРЕЖЬЯ СРЕДИЗЕМНОГО МОРЯ И З ЗАКАВКАЗЬЯ, А ТАКЖЕ СЕВЕРНУЮ АФРИКУ - 5 ВИДОВ (5,5%):
Все виды характерны для пустынных биотопов низинных долин и равнин. Из них Elaphria clara в Таджикистане встречается только в песках, не выше 400 м над ур. м. Остальные эремофилы поднимаются в пустынные низкогорья до 800 - 1000 м, a Pericyma albidentaria местами по сухим склонам доходит до 1700 м. Эта последняя распространена по всей пустынной зоне Средней Азии и достигает юго-востока Европейской части СССР Все остальные формы в Средней Азии не выходят за пределы ее западной части.
VIII ГРУППА. ВИДЫ, РАСПРОСТРАНЕННЫЕ В СРЕДНЕЙ АЗИИ И ЗАКАВКАЗЬЕ - 3 ВИДА (3,3%):

Все формы характерны для пустынных ландшафтов, как в Средней Азии, так и в Закавказье. Porphyrinia uniformis наиболее типична для тугайной растительности, Leucanitis sesquistria - для закрепленных песков, а Hadula sabulorum - для предгорий (два других вида в предгорьях не встречаются).
IX ГРУППА. ВИДЫ, РАСПРОСТРАНЕННЫЕ В СРЕДНЕЙ АЗИИ, НА ЮГО-ВОСТОКЕ ЕВРОПЕЙСКОЙ ЧАСТИ СССР ИЛИ ТАКЖЕ В ЮЖНОМ ПРИУРАЛЬЕ И НА СЕВЕРО-ВОСТОЧНОМ КАВКАЗЕ - 6 ВИДОВ (6,7%):

Ареалы одних видов, обозначенных звездочкой (*), не выходят за пределы Средней Азии и Юго-Востока Европейской части СССР, другие виды встречаются также в пустынной полосе Южного Приуралья и северо-восточной части Кавказа. Все формы характерны для пустынных биотопов и являются либо псаммофилами (Eriogaster henkei, Anumeta henkei и Leucanitis picta), либо эремофилами. При этом только Elaphria albina и Anumeta cestis поднимаются до нижней полосы среднегорий, остальные же виды не встречаются выше 400 м над ур. м.
X ГРУППА. ВИДЫ, РАСПРОСТРАНЕННЫЕ ОТ СРЕДНЕЙ АЗИИ И АФГАНИСТАНА ДО ЮЖНОЙ ЕВРОПЫ, ПЕРЕДНЕЙ И МАЛОЙ АЗИИ ИЛИ ТАКЖЕ ДО СЕВЕРНОЙ АФРИКИ - 15 ВИДОВ (16,5%):

Ареалы большей части видов, отмеченных звездочкой (*), простираются не только до Южной Европы, но и до Северной Африки. Три вида в Средней Азии представлены особыми подвидами (см. Список). В рассматриваемом комплексе имеются эремофилы, эврибионты, две тугайно-оазисные формы и равнинно-горные ксерофилы, но преобладают последние. Чисто низинных форм здесь совсем нет. Некоторые виды поднимаются в горы до 800 - 900 м (Cardepia sociabilis irrisor и Sideridis zeae indistincta), но большинство до 2000 - 2500 м, и только E. conspicua до 4000 м. Среди чешуекрылых, достигающих Северной Африки, преобладают наиболее теплолюбивые формы с наименьшими для данной группы пределами вертикального распространения.
XI ГРУППА. ФОРМЫ, ОЧЕНЬ ШИРОКО РАСПРОСТРАНЕННЫЕ В ПАЛЕАРКТИКЕ - 3 ВИДА (3,3%):

Ареалы охватывают очень большую территорию Палеарктики от Европы до Дальнего Востока, но Colias erate - преимущественно в южной части Палеарктики, а два других вида широко распространены и в средней полосе. Впрочем, и Melicleptria scutosa все же является преимущественно южнопалеарктической формой. Все виды более или менее ксерофильны; Xylina exoleta в горах идет до 2500 м, С. erate до снеговой линии и только М. scutosa в Северном Таджикистане пока не отмечена выше 700 м, а в Южном - выше 500.
XII ГРУППА. ГОЛАРКТЫ И КОСМОПОЛИТЫ - 5 ВИДОВ (5,5%):

Виды, отмеченные звездочкой (*), - космополиты. Для песчаной пустыни все эти формы не характерны, так как являются эврибионтами, причем С. lineata livornica - гемиксерофил, остальные - мезофилы. По вертикали Pieris rapae идет до 3000 м, но большинство до снеговой линии.
Мы видим, что господствующую по числу видов группу в фауне песков составляют эндемики Средней Азии - свыше 35%, притом бoльшинcтвo из них (24 вида, или более 26% всей фауны) эндемичны для ее западной части. Кроме того, ряд видов с более широкими ареалами представлен в Средней Азии или в ее западной части эндемичными подвидами. Псаммофилов здесь 20% от общего числа высших бабочек, или 59% всей группы эндемиков. Почти все остальные эндемичные виды, не будучи строгими псаммофилами, являются термо- и ксерофильными обитателями различных пустынных биотопов и распространены в низких жарких долинах, некоторые также в пустынных низкогорьях.
Таким образом, видовой эндемизм высших чешуекрылых даже самой далекой окраины Кара-Кумов, песков в низовьях Вахша, довольно высокий. Относительно высок и родовой эндемизм. В Кашка-Куме отмечаются представители 8 родов, ареалы которых не выходят за пределы Средней Азии, что составляет 12,4% от общего числа родов (65), представленных в вахшских песках. Число эндемичных родов и видов в песчаных пустынях всей Средней Азии, выше чем на их обедненной окраине, но, по-видимому, не достигает величины, указанной для некоторых семейств жуков. По данным О. Л. Крыжановского (1963), эндемичные виды среди среднеазиатских жужжелиц (Carabidae), карапузиков (Hesteridae), пластинчатоусых (Scarabaeidae), чернотелок (Tenebrionidae) и дровосеков (Cerambycidae) составляют 70% от общего их числа в Средней Азии, а эндемичные роды - 20%. Однако приведенные по жукам цифры касаются, во-первых, всей территории Средней Азии, в том числе и горной, а также Восточного Кавказа, во-вторых, жуки указанных семейств значительно менее подвижны, чем бабочки, и среди них, естественно, встречается большее число локальных форм. Тем не менее видовой и родовой эндемизм среди пустынных чешуекрылых Средней Азии, особенно ее западной части, достигает значительной величины и свидетельствует о большом своеобразии фауны песков (и вообще пустынь) и о наличии среднеазиатского пустынного центра формообразования.
Таким образом, материалы даже по высшим бабочкам, известным в большинстве своем как необычайно подвижные формы, подтверждают представления ряда зоологов и ботаников [А. П. Семенов - Тяньшанский (1936), В. Г. Гептнер (1945), О. Л. Крыжановский (1963) и А. Н. Краснов (1888), Е. П. Коровин (1962) и др.] о Средней Азии, во всяком случае о ее пустынях, как центре автохтонного формообразования.
Однако, характерным для Кашка-Кума является не только комплекс эндемичных форм. В ядро фауны песков Юго-Западного Таджикистана входят также и виды, общие для Средней Азии и окружающих ее стран пустынной зоны от Монголии на востоке до Белуджистана на юге, Передней Азии и Северной Африки - на юго-западе, Малой Азии и Закавказья па западе и до юго-востока Европейской части СССР, т. е. приуроченные к жарким низменностям восточной части древнего Средиземья (группы II, IV-IX групп). В числе форм, характерных для Вахшских песков, имеется ряд восточно-средиземноморских видов, распространенных в современную эпоху от восточного побережья Средиземного моря (VI группа) или также от южного его побережья в Северной Африке (VII группа) до Средней Азии и Афганистана.
Однако, уже более широко распространенные средиземноморские виды, от Средней Азии до Южной Европы или также до Северной Африки (X группа), в песках попадаются в небольшом числе, да и то на окраине, и для них не типичны. В совокупности средиземноморцев (восточных и широко распространенных) насчитывается лишь 25 видов, т. е. 27,7% всей фауны высших; из них только меньшая часть (2/5) входит в ядро фауны.
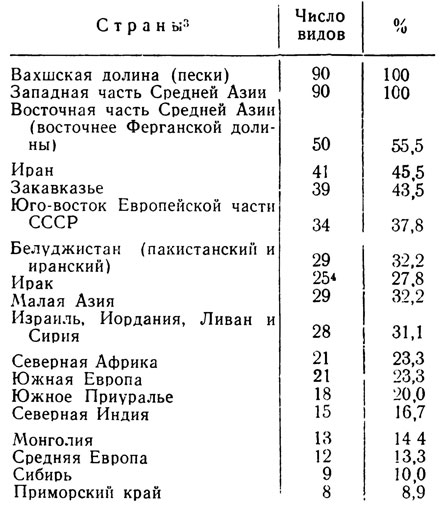
Таблица
3(Афганистан не включен в таблицу, так как в самой северной части его (по Аму-Дарье) встречаются все виды, имеющиеся в Вахшской долине, но центральные и южные его районы изучены крайне слабо)
4(В Ираке, вероятно, встречается больше общих видов, но он слабее изучен, чем Иран или страны западного побережья Средиземного моря.)
Наибольшее число общих со Средней Азией видов оказывается в странах, лежащих полукругом к югу, юго-западу и северо-востоку от нее: в Афганистане, Иране, Закавказье и на юго-востоке Европейской части СССР, причем при движении с юго-востока на северо-запад в этом ряду наблюдается убывание количества форм, а в самой Средней Азии основная масса этих общих видов сосредоточена в ее западной части (см. табл. 2). Затем по числу общих форм в убывающем порядке следуют районы второго полукольца: Белуджистан, Ирак, страны восточного побережья Средиземного моря, Малая Азия. Еще меньше видов, общих для северной Африки, Южной Европы, Южного Приуралья и совсем мало в Сибири и Приморском крае. Так что, только формы, не выходящие за пределы Средней Азии, Афганистана, Ирана и Ирака, составляют половину всей фауны. В экологическое же ядро фауны вахшских песков входит 57 видов, или 63,3% от общего числа высших чешуекрылых. Но если не учитывать частоту встречаемости форм, их обилие в песках и принять во внимание лишь географию видов (I, II, IV - IX групп) как наиболее характерных для пустынь вообще, а не только песчаных, то в Кашка-Куме в этом комплексе насчитывается 66 видов, или 73,3% всей фауны высших. В основной своей массе он состоит из псаммофилов и более широких по специализации эремофилов и включает очень небольшое число равнинно-горных ксерофилов. Из приведенных цифр ясно, что фауна песчаных пустынь Средней Азии имеет прямые генетические связи, в первую очередь, с ближайшими странами, лежащими от нее к югу, с Афганистаном и Ираном, затем с остальными, составляющими указанные два полукольца до восточного побережья Средиземного моря и Нижнего Заволжья включительно. При этом выявляется совершенно определенная связь именно с жарки ми пустынями этих стран, но отнюдь не с фауной этих областей в целом. Связи с Монголией и Индией несравненно слабее; но фауна первой, особенно ее песчаных пустынь исследована еще совершенно недостаточно и можно ожидать, что фаунистическое родство пустынь Средней Азии и Монголии более близкое, чем представляется в настоящее время. Что касается Западной Индии, точнее ее пустыни Тар, то лепидоптерофауна последней, несомненно, палеарктическая и представлена рядом типичных пустынных форм, близких к нашим (например, виды p. Anumeta, в том числе и псаммофилы), но там почти нет видов, общих со Средней Азии характерных пустынных, хотя встречается ряд общих видов экологически более пластичных.
Что же представляют собой несколько остальных, приведенных выше, групп чешуекрылых, не составляющих ядра фауны? Это - формы, широко распространенные от Средней Азии до Южной Европы или также до Северной Африки (т. е. широкие средиземноморцы - X группа) или еще шире - также в Сибири и Северной Америке (субпанпалеаркты и голаркты - группы XI и XII) и даже по всему земному шару (космополиты - группа XII). Всего в этих группах содержится лишь 23 вида (25,5%). Но и в этом комплексе большинство форм, будучи южно-палеарктическими термофилами, в той или иной степени ксерофильны, хотя они встречаются не только в пустынных биотопах; из них 9 видов в условиях Средней Азии являются преимущественно пустынными или полупустынными элементами (Autophila cerealis, Microloxia herbaria, Acidalia beckeraria, Oria musculosa laeta, Cardepia sociabilis irrisor. Euchloe belia pulverata, Chloridea nubigera, Melicleptria scutosa, Rhodometra sacraria). Из всей фауны песков только Papilio machaon, Pieris rapae, Pyrameis cardui и Scotogramma trifoli являются мезофилами. Эти четыре вида удерживаются местами в песках лишь благодаря своей исключительной пластичности, приспособленности к самым различным экологическим условиям, но и они в песках встречаются лишь весной и в самом начале лета.
Остается сказать о третьей группе. В Вахшской долине индийские горные элементы представлены очень слабо - двумя горно-долинными видами. Только один из них - Neoris stoliczkana schencki - по низкогорьям и небольшим горкам (в данном случае через г. Буритау) проникает в пески, но только на окраину их, где встречается одно из кормовых растений - парнолистник Гончарова. Значительно больше видов этой группы в среднегорной полосе; в песках же на нее приходится лишь 1,1 % от общего числа высших бабочек.
Таким образом, фауна песков состоит в основном из сильно специализированных пустынных форм, многие из которых приспособлены для жизни только в песках. Распространение большинства характерных видов ограничено, как сказано, пустынной зоной Средней Азии и странами, лежащими от нее на юг и юго-запад. Здесь совершенно очевидна тесная связь географического распространения и экологической приуроченности видов. Но экологическая приуроченность является важнейшим фактором, определяющим ареал вида. Исторический фактор, конечно также отражается на современном распространении форм, однако развитие вида в далеком прошлом нельзя представлять себе изолированно от определенных экологических условий. Вот почему характерные формы для того или иного биотопа часто имеют и сходные ареалы. Но внешнее сходство ареалов видов, обитающих в несходных ландшафтах, не всегда свидетельствует об общности происхождения форм. Внешнее подобие ареалов в таких случаях может определяться лишь особенностями распределения ландшафтов. В Передней, Средней и Центральной Азии, например, рядом с горными цепями, часто параллельно им, находятся низкие жаркие долины, пустыни и т. д. Естественно, что грубо вычерченные, без учета экологической приуроченности, ареалы видов, обитающих в разных ландшафтах и на различных высотах, в иных случаях оказываются внешне сходными. К тому же, границы обширных ареалов (например, панпалеарктических) нередко определяются опять-таки климатическими факторами, общими для определенной большой территории, хотя внутри ее виды размещаются по очень, казалось бы, несходным экологически зонам и поясам. Исторический фактор в современном распределении видов, несомненно, имеет значение, однако он не является единственным и, более того, не может быть главным, определяющим. Каковы бы ни были исторические условия развития и перемещения вида в прошлом, он занимает в каждый период своей истории только тот биотип (или группу биотопов), в котором удовлетворяются его потребности. В противном случае он не мог бы существовать. Поэтому при рассмотрении видов с действительно, а не только внешне, сходными ареалами, обнаруживаются и другие особенности, общие данной группе видов, в первую очередь, сходство в экологической приуроченности, проявляющейся, в частности, в определенном распределении по горизонтали и вертикали.
В таком относительно однородном биотопе, как пески Кашка-Кум, большинство его характерных специализированных видов приурочено к более или менее обширной зоне пустынь древнего Средиземноморья. Здесь влияние истории этой части земной поверхности в третичное и четвертичное время на распространение видов очевидно. Но также очевидно, что определяющим фактором оказалось именно образование зоны пустынь, в которой и формировались термоксерофильные формы и их ареалы. В горах же Средней Азии и соседних стран распределение видов произошло иначе, в соответствии с климатическими условиями тех или иных поясов.
Как сказано, современное географическое родство видов, имеющих даже сходные или почти тождественные ареалы, не всегда свидетельствует о происхождении фауны того или иного биотопа из единого центра. Географическое родство в иных случаях менее древне, чем экологическая общность видов. Глубокие генетические корни у некоторых форм с однотипными ареалами, даже из одного и того же биотопа, уходят в различных географических направлениях, так что далекие предки этих видов (в иных случаях экологически близкие, как и в современную эпоху) обитали в весьма отдаленных друг от друга местностях земного шара. Рассмотрим для примера IV группу видов Кашка-Кума, в которую включены формы, распространенные в Средней Азии, Афганистане и Восточной части Ирана, доходящие на юг до Белуджистана. У половины видов этой группы ареалы в Средней Азии охватывают почти всю пустынную зону Средней Азии или местами даже выходят за ее пределы. Ареалы других, более локальных видов в Средней Азии занимают лишь ее западную часть. У этих последних, наиболее специализированных видов, типичных эремофилов или псаммофилов, ареалы очень сходны. Однако, если три из четырех видов (Armada dentata, Leucanitis sinuosa и Atomorpha hedemanni) по происхождению являются палеарктами, то четвертый вид - Chilena sordida - принадлежит к роду, явно тяготеющему к эфиопской фауне. Из 9 палеарктических видов рода Chilena 7 обитают только в Северной Африке. Наибольшее число (10) видов рода сосредоточено в Эфиопской области, причем все формы (также 10 видов) очень близкого к Chilena рода Beralade известны лишь из Эфиопской области и многие из них характерны для засушливых районов Южной Африки.
В VI группе видов, распространенных от Средней Азии и Афганистана до Передней Азии и Закавказья, все формы, в том числе и Chondrostega pastrana, явно пустынные, восточно средиземноморские. Подавляющее большинство видов (15) рода Chondrostega сосредоточено в восточных и южных странах Средиземноморья (в частности, в Северной Африке) и только некоторые отмечены в Иране и Средней Азии. Но два вида этого рода обитают в пустынных областях Южной Африки, причем там же обнаружен и 1 вид из ближайшего (южноафриканского) монотипичного рода Chondrostegoides. Можно полагать, что относительное обилие видов p. Chondrostega в странах Средиземноморья свидетельствует о возникновении здесь вторичного центра видообразования.
М. Г. Попов (1927), кажется, первый из советских ботаников обратил внимание на связи пустынных флор Палеарктики и Южной Африки, разделенных в современную эпоху широкой зоной тропиков. Он считал, что центр возникновения и развития первичной пустынной покрытосеменной флоры земного шара находился на материке Гондвана (в меловое или даже юрское время). После разрушения Гондваны последующее развитие пустынной флоры продолжалось в Капской области и в проблематичной Лемурии. Из Южной Африки в начале палеогена или еще в верхнемеловое время происходила миграция флоры на север этого материка и далее в Переднюю и Среднюю Азию, где возникали вторичные пустынные центры формообразования. Этой концепции придерживаются многие другие ботаники и в настоящее время. Однако другие авторы (Ильин, 1936, 1946; Невский, 1937) придают пустынной флоре Средней Азии несравненно большее значение, считая ее самобытной и древней, как и гондванская флора.
Что касается зоологических объектов, то на материале чешуекрылых, насколько известно, этот вопрос не обсуждался, а между тем приведенные выше примеры дают основания для аналогичных суждений с теми, что были высказаны ботаниками. Во всяком случае, с одной стороны, очевидна связь современных пустынных фаун чешуекрылых Палеарктики и Эфиопской области, фаун, разделенных широким поясом тропических лесов экваториальной Африки; с другой стороны, это свидетельствует о прямых связях пустынь в далеком прошлом, вероятно, еще в дотретичное время. Но эти связи не многочисленны, и хотя они свидетельствуют о миграциях пустынных элементов в верхнемеловое время или в нижнем палеогене, однако в настоящую эпоху состав пустынных фаун Южной Африки и Палеарктики все же настолько различен (несмотря на некоторые родственные корни), что приходится признать их необычайную самобытность, большую древность и обособленность. Наконец, наши примеры подтверждают сказанное выше о том, что даже в однородном биотопе виды со сходными ареалами могут быть различного происхождения. Это не значит, что буквально все или многие виды биотопа с данным типом ареала имеют различное по отношению друг к другу географическое происхождение. Большинство форм, имеющих сходные ареалы, даже не будучи родственными в систематическом отношении, оказываются происходящими из единого центра формообразования или географически близких центров в относительно недалеком прошлом. Но среди них нередко отмечаются виды, глубокими корнями связанные явно с другими регионами.
Если последнее относится к экологически близким формам из одного и того же биотопа, то в еще большей степени это касается фауны какого-либо участка суши в целом, фауны экологически крайне пестрой и тем более включающей представителей ряда вертикальных поясов. Поэтому, если можно еще говорить об общности происхождения комплекса форм отдельного биотопа или ряда близких биотопов (например, пустынных), то нельзя эту же мысль отстаивать априорно в отношении всей фауны территории с самыми разнообразными ландшафтами на различных высотах над уровнем моря с резко отличными климатическими, почвенными и гидрологическими условиями.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://butterflylib.ru/ 'Чешуекрылые, бабочки, мотыльки, моли'