Экологические группировки и вертикальное распределение чешуекрылых песчаной пустыни
Общим вопросам экологии каких-либо комплексов чешуекрылых уделялось до сих пор очень мало внимания, а песчаная фауна в этом отношении вообще не затрагивалась. Данная работа представляет первую попытку вычленения экологических группировок чешуекрылых песков, но в этом имеются свои трудности, так как принципы выделения экологических группировок пустынь не разработаны, а биология многих видов остается неизвестной. Фауна не только пустынь вообще, но и песчаной пустыни, представлена экологически неоднородными элементами. Это обнаруживается по распределению чешуекрылых в различных пустынных биотопах и по частоте встречаемости тех или иных видов. Казалось бы естественным выделить группировки по степени приспособленности чешуекрылых к тем или иным факторам, характерным для биотопа (к высоким температурам, сухости воздуха, сыпучести грунта и т. п.). Однако одни адаптации организмов могут быть относительно легко, так сказать, обозримы и учтены (например, многие морфологические приспособления организмов к жизни в пустыни), другие скрыты для поверхностного исследования (например, большинство физиологических адаптаций), и без специальной экспериментальной работы остаются недоступными полевому исследованию. С другой стороны адаптации имеются на всех фазах развития организма и соответствуют определенному сезону года. У многих пустынных чешуекрылых куколка приспособлена к длительным исключительно высоким летним температурам, тогда как у гусеницы, развивающейся весной, отсутствуют какие-либо адаптации к ксеротермическим условиям, так как она в этом не нуждается, имаго также может быть не приспособлено к выживанию в крайне сухой воздушной среде при высоких температурах по той простой причине, что вылет из куколки происходит осенью и к тому же бабочки днем скрываются в укромных затененных местах. В целом же вид оказывается хорошо приспособленным к жизни в песчаной пустыне, так как его жизненный цикл и распределение фаз развития во времени строго адаптированы к сезонному ритму года.
Поэтому нельзя выделить экологические группировки по адаптациям организмов к отдельным факторам среды или по адаптациям отдельных фаз развития. Приходится учитывать всю совокупность самых разнообразных адаптивных свойств организма. К сожалению, биологию многих пустынных видов мы вообще не знаем, других - слишком поверхностно, а физиология всех форм пока остается не затронутой исследованиями. Но в конечном счете адаптивные особенности организмов проявляются в их экологической приуроченности у одних очень строгой, у других более широкой, в чем выражается степень специализации или лабильности организмов. Однако для вычленения группировок на основании экологической приуроченности, конечно, требуется достаточный опыт полевых исследований в данном районе и знание фауны различных биотопов, систематичные многолетние сборы, как базис для работы.
Выделив основные экологические группы, из которых состоит комплекс чешуекрылых данного биотопа, внутри их уже можно подразделить по месту обитания, образу жизни, кормовым растениям или на основании адаптации к каким-либо другим определенным факторам среды.
По экологической приуроченности чешуекрылых песков Юго-Западного Таджикистана можно разбить на 5 основных групп: I - псаммофилы; II - эремофилы; III - тугайные и оазисные ксерофилы; IV - равнинно-горные ксерофилы; V - эврибионты.
В число псаммофилов включены крайне специализированные формы, обитающие в пределах Таджикистана только в песках. Вне песков, если некоторые иногда и попадаются в имагинальной фазе, то лишь как залетные:

Преимагинальные фазы и жизнь большинства перечисленных форм остаются не известными науке. Мне удалось изучить в основных чертах биологию только 6 видов: Praephilotes antracias, Eriogaster henkei, Lemonia tancrei, Anumeta cestina, A. spilota, Leucanitis picta. Формы рассматриваемой группы или связаны с псаммофитными кустарниками (монофаги), или (большинство) с эфемерово-эфемероидной растительностью (монофаги и олигофаги). Жизнь в песках у одних видов (Praephilotes antracias, Eriogaster henkei, Anumeta spilota, An. cestina) обусловлена, быть может, только их монофагией, у других (например, Lemonia tancrei), связанных с экологически более широко распространенными растениями (эфемерами), определяется какими-то другими факторами, пока нам не известными. Большая часть их моновольтинна, что часто связано с питанием эфемерами. Лишь 1/5 видового состава этой группы развивается за год в нескольких поколениях (Anumeta spilota, An. cestina, Leucanitis sesquistria, L. picta, L. sinuosa). Из 26 видов 16, т. е. более трех пятых, эндемичны для Средней Азии (в том числе 11 эндемов из западной ее части). Девять других видов имеют несколько более широкое распространение в песках пустынной зоны: в Средней Азии, Афганистане, Иране, Ираке и только один вид доходит до Северной Африки (Elaphria clara). Высотное распространение всех форм ограничено равнинами и низкими жаркими долинами и не превышает 400 м над ур. м.
Все остальные виды, отмеченные в песках, но не входящие в рассмотренную группу, являются общими для песчаной пустыни и других (одного или нескольких) биотопов долины. Среди них самой обширной является следующая группа.
В этот комплекс тоже входят гиперксерофильные виды, характерные для различных жарких пустынных безводных биотопов равнины, низких долин и нижней полосы низкогорий. Не вошли сюда только псаммофильные формы, составившие предыдущую группу. Термин "эремофилы" означает вообще пустынные формы, но здесь он использован в более узком значении.
По кормовым растениям эремофилов можно распределить в 3 подгруппы.

* (Биология многих видов остается не известной, но можно с уверенностью сказать, что виды этой подгруппы питаются весенне-вегетирующими растениями, так как встречаются и в местностях, где нет кустарников или многолетников.)
Данная подгруппа по числу видов почти вдвое больше двух остальных и равна группе псаммофилов. По видовому составу она близка к аналогичному комплексу эремофильных форм предгорной фауны низовья Вахша. Виды, отмеченные звездочкой (*), в Вахшской долине встречаются преимущественно в песках; обозначенные двумя звездочками (**) в песках сравнительно многочисленны, но в некоторых других биотопах долины могут встречаться в таком же или еще большем количестве. В общем, отмеченные звездочками виды для вахшских песков очень характерны и входят в экологическое ядро песчаной фауны долины (эти обозначения использованы и в других списках главы).
Гусеницы питаются однолетними травянистыми растениями. Цикл развития чешуекрылых теснейшим образом связан с сезонным развитием кормовых растений: все виды за год дают только одно поколение, активная фаза гусеницы приходится на период весенней вегетации кормовых растений, фаза куколки и куколочная летняя диапауза у многих форм - на время летней жары, когда эфемеры, закончив развитие, желтеют и отмирают. Лёт бабочек происходит осенью или, у большинства, весной (у некоторых видов в начале лета) следующего года.
Почти половину этой подгруппы (11 видов) составляют среднеазиатские эндемики, а ареалы такого же числа других видов не выходят за пределы Средней Азии, Афганистана, Ирана, Ирака, стран Палестины, Малой Азии, Закавказья и пустынных районов низовий Волги. И только 3 вида (Agrotis lasserei, Oria musculosa и Armada panaceorum) распространены значительно шире: до Южной Европы или Северной Африки, а Scithocentropus scripturosa - до Монголии. По вертикали формы рассматриваемого эремофильного комплекса распространены следующим образом: 10 видов не поднимаются выше 450 - 500 м над ур. м., 13 - идут не выше 700 - 800 м, два - до 1100 - 1300 м (Oria musculosa, Elaphria asymmetrica), только два местами достигают 2000 - 2 300 м (P. tesselata, S. apotheina). Таким образом, основная масса видов обитает на небольших высотах и характерна для долин и пустынных низкогорий, не поднимаясь выше 800 м над ур. м.

Биология большинства видов этой подгруппы известна. Виды, кормовые растения которых не найдены, отнесены сюда на основании характера лёта бабочек (поливольтинные) и особенности распределения по биотопам в долине и других местностях. Подавляющее большинство форм поливольтинны, что имеет прямую связь с питанием, многолетниками, вегетирующими в течение всего лета. Приспособленность к крайне ксеротермическим условиям пустыни у большинства видов проявляется в способности переносить исключительную сухость воздуха и очень высокие летние температуры почти во всех фазах. Некоторые (С. boryphora) имеют летнюю диапаузу в фазе куколки и дают лишь две генерации за год, и лёт бабочек происходит при умеренных температурах весной и осенью. Половина видов питается верблюжьей колючкой - Alhagi canescens. Все они монофаги (Apharitis epargyros, Chilena sordida, Pericyma albidentaria, Leucanitis flexuosa. Rhynchodontodes ravalis). Некоторые формы живут на полыни (Melicleptria scutosa и, вероятно, Cucullia boryphora), Chloridea nubigera - на бобовых.
Большинство форм рассматриваемого комплекса в песках немногочисленно и не характерно для них, что имеет прямую связь с распределением в долине их кормовых растений. Только Leucan. flexuosa и Cucullia boryphora в Вахшских песках обычны.
Впрочем, первая, как и большинство остальных, все же более типична для тугаев и сорной растительности в поливной полосе земель, где верблюжья колючка достигает пышного развития и часто образует обширные заросли.
Географическое распространение некоторых видов ограничено в основном Средней Азией и сопредельными с нею местностями (Apharitis epargyros, Chilena sordida, Cucullia boryphora). Эндемиков всего два (Porphirynia gratiosa и Aridargricola rufulus), т. е. 1/5 от общего числа форм группы. Широко распространенные формы (до Южной Европы или Северной Африки) составляют половину общего числа, но из них лишь Melicleptria scutosa распространена в Палеарктике очень широко (хотя в условиях Средней Азии, в частности и в Вахшской долине, она характерна для пустынных биоценозов).
Большинство видов приурочено к полосе от низких долин через предгорья до низкогорий, в пределах 300 - 800 м над ур. м. Только Porphyrinia gratiosa, Aridagricola rufulus и Melicleptria scutosa в Таджикистане не отмечены выше 400 - 650 м. Три вида проникают в среднегорья и местами, где есть подходящие условия, доходят до 1200 м (Rhynchodontode.s ravalis), 1700 м (Pericyma albidentaria) и даже выше (С. boryphora), но у верхнего предела своего распространения редки.
К этой подгруппе, представленной более обильно в предгорьях, в песках принадлежит лишь 3 вида, которые для песков не характерны и чаще встречаются в тугаях и некоторых оазисах:

Одноядные формы, питающиеся такими псаммофильными кустарниками, как джузгун, саксаул, солянка Рихтера и не выходящие за пределы песчаной пустыни, отмечены выше (псаммофилы). Рассматриваемые эремофильные кустарниковые формы по кормовым связям являются олигофагами.
Cardepia sociabilis irrisor живет в песках на белом саксауле (Haloxylon persicum Bge.), а в других биотопах - также на саксаульчике (p. Hammada), и, очевидно, на различных тамариксах (p. Tamarix). К этой же категории можно была бы отнести и Pseudohadena siri, отмеченную на саксауле в Туркмении (Даричева, 1960а), но в вахшских песках этот вид не найден, хотя встречается в других биотопах. Cardepia sociabilis характерен для низменных пустынных, часто солончаковых, биотопов. В горах не встречается.
Elaphria albina, так же как и предыдущий вид, за год дает несколько поколений. Распространена как в долинах, так местами и в среднегорьях. Кормовым растением в Вахшской долине, вероятно, служит тамарикс; в других местах Средней Азии отмечена на саксауле.
Hypermnester helios за год дает одно поколение, питается видами парнолистника (p. Zygophyllum), не только из группы травянистых видов (Z. fabago и др.), но также и кустарниковыми формами (в низовье Вахта на Z. Gontscharovii). Встречается в низкогорьях до высоты 800 - 850 м.
В рассматриваемом комплексе чешуекрылых нет эндемиков, но ареал Hypermnestra helios ограничен лишь Средней Азией, Афганистаном и Ираном. Оба других вида идут далеко на запад (до Южной Европы), характерны для пустынных биотопов.
Таким образом, основная масса эремофильного комплекса состоит из видов, питающихся весенними, эфемеровыми растениями и в связи с этим, развивающихся за год в одном поколении. Такие виды составляют 66,7% группы, или 29% всей фауны высших бабочек песков. Форм, живущих на многолетних травах, вегетирующих в течение всего лета немногим более 1/4 (25,6%), а питающихся кустарниками лишь около 8% (7,7) от общего числа группы. Виды этих двух последних подгрупп более многочисленны в предгорьях, в песках же они занимают скромное место. Почти все они, кроме Hypermnester helios, поливольтинны.
По вертикали большинство эремофилов распространено в больших пределах, чем псаммофильные формы. Видов, не поднимающихся выше 500 м над ур. моря, лишь около 25,6% от общего числа этой группы, тогда как идущих до 700 - 900 м - более половины (свыше 56%); в то же время 1100 - 1200 м достигает 4 вида (10,3%) и лишь 3 вида по сухим склонам местами поднимаются в среднегорной полосе до 2000 м (7,7%).
Значительную часть эремофилов составляют среднеазиатские эндемики - 14 видов, или 35,9% всей группы, 7 видов (17,9%) распространены в Средней Азии, Афганистане, Иране и Ираке; кроме того у 11 видов (свыше 28%) ареалы находятся в пределах Средней Азии и стран, расположенных к югу и западу от нее до Северной Африки, Закавказья и Юго-Востока Европейской части СССР и охватывают большую или меньшую часть этой территории. Наконец, видов, распространенных от среднеазиатских республик до Северной Африки и также до Юго-Западной Европы всего 5 (12,8%), а очень широко распространенных в Палеарктике форм только одна. Голарктов и космополитов в группе эремофилов нет совсем.
Эремофилы приурочены повсюду к пустынным биотопам. Многие из них характерны для песков, хотя распространены не только в этом биотопе. Многие проникают в поймы рек или на поливные возделываемые земли, а некоторые находят в этой обстановке даже лучшие условия для развития; однако и там они держатся преимущественно на сухих опустыненных участках.
а)тугайные формы (2 вида):
Sideridis zeae indistinct Porphyrinia uniformis
б)тугайно-оазисные (1 вид):
Rhodometra sacraria
Первые два вида приурочены преимущественно к тугайной пойме, для песков не характерны и встречаются там в очень небольшом числе. Они найдены однако не только в низовьях Вахша, но также на участках бугристых песков по Сурхан-Дарье около г. Джар-Курган в Южном Узбекистане, a Sideridis zeae indistincta и в Гиссарской долине, где однако очень редка; в горах отсутствуют, а в долинах не отмечены выше 500 - 800 м над ур. м. Если S. zeae распространена широко (до Южной Европы и Северной Африки), то Porphyrinia uniformis - лишь от пустынных районов Закавказья до Средней Азии. Rhodometra sacraria в песках довольно обычна, но в Таджикистане встречается в значительно большем числе в культурном ландшафте; нередка также и в тугаях. Как и предыдущие виды, поливольтинна. Ареал обширный: по всей Средней Азии, до Белуджистана и Западной Индии, Северной Африки и Южной Европы. В горах Таджикистана отмечена до высоты 1100 м
В данную группу входят ксерофильные (но не гиперксерофильные) формы, обитающие как в низких долинах и на равнинах, так и в горах. При этом очень резкого численного преобладания на какой-либо высоте в пределах 300 - 2000 м над ур. моря обычно не наблюдается. В списке двумя звездочками (**) отмечены формы, сравнительно обычные в песках, но из менее многочисленные и в некоторых других биотопах, в том числе и в горных. Среди равнинно-горных ксерофилов одни виды питаются эфемеровыми растениями, другие многолетниками, третьи кустарниками. На основании приуроченности к определенной категории растений они подразделены в три подгруппы.


Neoris stoliczkana schencki
В рассматриваемой группе, численно сильно уступающей комплексу эремофилов, как и среди последних, основной контингент, т. е. более половины форм, питается травянистыми эфемеровыми и эфемероидными растениями (1-я подгруппа). Численное преобладание форм, питающихся весенними растениями в этой группе, как и среди псаммофилов и эремофилов, имеет прямую связь с господством эфемеров и в конечном счете с пустынным климатическим режимом биотопа. Будучи ксерофилами, эти виды однако более лабильны, чем типичные пустынные формы, псаммофилы или эремофилы. Пластичность их проявляется в большем диапазоне вертикального распространения и, как правило, в более обширном ареале, а также, в приуроченности не только к пустынным ландшафтам, но и к другим биотопам с более или менее ксеротермическим режимом. В горах они обычно встречаются на сухих хорошо освещенных и прогреваемых солнцем склонах. Виды, живущие на эфемерах, развиваются в одном поколении. По вертикали почти все достигают 2000 (Euchloe belia pulverata) - 2500 м (Margelana versicolor, Ogygia flammatra), а некоторые доходят до 3000 м над ур. м. (Satyrus enervata, Neolycaena tengstroemi, Lasiocampa eversmanni), и только Cucullia chamomillae не отмечена выше 1100 м. Формы, питающиеся летневегетирующими многолетниками (поливольтинные и моновольтинные), составляют лишь 1/3 группы. У видов этой (2-й) подгруппы ксерофильные свойства выражены, пожалуй, менее отчетливо. Если в 1-й подгруппе ряд видов довольно многочислен в песках (Euchloe belia pulverata, Lasiocampa eversmanni, Margelana versicolor), то представители 2-й подгруппы там встречаются не так часто, как, например, в оазисах, тугаях, или в горах. Верхний предел вертикального распределения видов 2-й подгруппы ограничен - 2000 - 3000 м. Однако горизонтальное распространение их обширно: ареал всех четырех видов охватывает и Южную Европу, тогда как более половины видов, живущих на эфемеровой растительности, в своем распространении ограничены в основном или только территорией Средней Азии, Афганистана и Ирана (Satyrus enervata, Neolucaena tengstromi, Margelana versicolor) или же на западе доходят лишь до Малой Азии, Закавказья и юго-востока Европейской части СССР (Lasiocampa eversmanni).
Наконец, из чешуекрылых, питающихся древесно-кустарниковыми породами, в рассматриваемой группе только один вид - моновольтинный и преимущественно горный, достигающий местами 3500 м над ур. м., и для песков не характерный. По горизонтальному распространению и происхождению он тяготеет к Ориентальной области.
Обобщая сказанное о равнинно-горных ксерофилах, следует еще раз подчеркнуть, что большинство видов развивается в одном поколении за год и связано с травянистыми весенними однолетниками, распространено по вертикали в довольно большом диапазоне - от 300 до 2000 - 3000 м над ур. м., а по горизонтали (2/3 видов) до Южной Европы. Эндемиков нет совсем. Почти все формы олигофаги или полифаги.
Здесь объединены гемиксерофильные и мезофильные формы, распространенные в самых разнообразных биотопах и многочисленные как в низких долинах, так и в горах Средней Азии.

Ксерофильные особенности видов этого комплекса выражены относительно слабее, чем у представителей всех других рассмотренных групп. В песках перечисленные формы, кроме Celerio lineata livornica и Xylina exoleta impudica, немногочисленны. В низких долинах они наиболее характерны для культурного ландшафта, но отнюдь не для безводных биотонов; в горных районах встречаются как на сухих южных склонах, так и на более влажных, с мезсфитной, луговой, растительностью, но преимущественно на открытых солнечных местах. При этом верхняя граница распространения у одних лежит на высоте - 2000 - 2500 м (Porphyrinia parva, Р ostrinа, Xylina exoleta), у других у снеговой линии. Ареалы их охватывают обширную территорию от Средней Азии до Южной Европы или еще большую (Colias erate, Celerio lineata livornica). Все виды многоядны и, кроме Euxoa conspicua, за год дают несколько поколений.
К числу мезофилов отнесены формы, характерные для орошаемых земель, участков с мезофитной растительностью (люцерновые поля, сады, приарычная сорная растительность). В горах они также наиболее многочисленны на более влажных склонах и в поймах рек. Только Papilio machaon типичен для открытых солнечных пространств и склонов, особенно обычен на вершинах возвышенностей и отдельных горок. В Вахшской долине он найден только на вершине Песчаного перевала и на г. Буритау. Все четыре вида распространены очень широко: по всей Голарктике или даже всесветно (Pyrameis cardui). В горах Средней Азии одни доходят до высоты в 3000 м (Pieris rapae) или еще выше, до снеговой линии (остальные). Все формы многоядные и поливольтинные. В песках развиваются только в первой половине лета (одно - два поколения), а затем либо диапаузируют (вероятно, Pap. machaon), либо мигрируют в тугайную пойму или на поливные земли, где имеются кормовые растения, вегетирующие до осени. Развитие их происходит обычно на мезофитных травянистых растениях, реже и на ксерофитах.
В общем эврибионтные виды слабо ксерофильны или мезофильны, преимущественно поливольтинные, все очень широко распространены как горизонтально, так и вертикально, причем верхняя граница распространения у большинства форм лежит не ниже 3000 м над ур. м. (чаще выше). Все рассматриваемые виды многоядны, питаются травянистыми растениями, однолетниками или многолетниками, или и теми и другими.
Рассмотрев отдельные экологические группировки песков Вахшской долины, можно сделать следующие выводы. Характер фауны чешуекрылых песков, и вообще любого биотопа, определяется климатическими, почвенными и гидрологическими особенностями биотопа. Эти физические факторы во всем их сезонном многообразии оказывают как непосредственное влияние на видовой состав чешуекрылых в процессе исторического формирования пустынной фауны, так и через пустынную растительность, с которой теснейшим образом связано развитие насекомых, а также через представителей всего остального биоценоза и в особенности хищников и паразитов. В песках выживают лишь виды, более или менее способные переносить крайне высокие летние температуры, исключительную сухость воздуха, приспособленные к сыпучим свойствам и особому гидрологическому режиму грунта.
Подавляющее большинство видов (95,5%) чешуекрылых песчаной пустыни является ксерофилами. Мезофилы (4,5%) для рассматриваемого биотопа не характерны, встречаются в очень небольшом количестве и почти все только весной или в начале лета. Почти 3/4 видового состава представлено термо-гиперксерофильными формами, в число которых входят высоко специализированные виды, приспособленные к жизни в песчаной пустыне, псаммофилы, и чешуекрылые, характерные для различных других пустынных, жарких и безводных, ландшафтов - эремофилы. Однако из группы эремофилов в песках не все виды многочисленны. Ядро фауны песков Вахшской долины составляют как формы, приуроченные исключительно к пескам, так и другие гиперксерофильные и ксерофильные, достаточно многочисленные в этом биотопе. В этот основной контингент входят, таким образом, все псаммофильные формы (26 видов), 25 эремофилов (13 видов, в Вахшской долине встречающихся преимущественно в песках, и 12 других, более или менее многочисленных в песках), а также некоторые сравнительно многочисленные представители группы равнинно-горных ксерофилов (Euchloe belia pulverata, Neolycaena tengstroemi,* Lasiocampa eversmanni atritta, Margelana versicolor) и очень немногие полуксерофильные эврибионты (Celerio lineata livornica, Xylina exoleta impudica). Всего экологическое ядро составляют 57 видов (63,3% от общего числа высших бабочек).
* (Neolycaena tengstroemi в Вахшских песках редка, но вообще она характерна именно для песков на равнине и для некоторых биотопов в горах, В Вахшской долине она найдена только в Кашка-Куме.)
Основная масса форм развивается на весенних, эфемеровых и эфемероидных, растениях. Это имеет место во всех группировках, кроме тугайных видов и эврибионтов, питание которых происходит на многолетних травянистых растениях (у первых) или как на эфемерах, так и на многолетниках (у вторых). При этом в группе псаммофилов на ксерофильных кустарниках развивается 1/4 - 1/3 всего видового состава, остальные формы все или почти все - на эфемерах. Около 3/4 эремофилов питаются травянистыми весенними растениями, около 1/4 - многолетниками и менее 1/10 - кустарниками. В комплексе равнинно-горных ксерофилов (четвертая группа) на эфемерах развивается немногим более половины видов, на многолетниках - 1/3 и на кустарниках - 1/10. Такое распределение в экологических группах объясняется характером самой растительности, с которой теснейшим образом связан видовой состав бабочек: в песчаной пустыне имеются заросли пескоукрепительных кустарников и их значение как кормовых растений весьма заметно; в эфемеровой пустыне или в других типах пустынь, где кустарников и многолетников нет или очень мало, развитие громадного большинства чешуекрылых происходит за счет эфемеровой и эфемероидной растительности. В группе эврибионтов в питании большинства форм (в долине) в том или ином поколении имеют значение культурные растения, в том числе и многолетние (люцерна, клевер, капуста, редька, хрен и др.).
Две трети видов развиваются в одном поколении. Моновольтинизм подавляющего большинства форм связан, с одной стороны, непосредственно с исключительно жестким ксеротермическим режимом пустыни, с другой стороны, с господством весенних однолетников. Все чешуекрылые, живущие за счет весенних травянистых растений, моновольтинны. Впрочем, одну генерацию в году дает также и ряд видов, питающихся многолетниками и кустарниками. Во многих случаях моновольтинизм - исторически выработавшаяся особенность всего рода или группы близких видов. Возможно, подобные группы близких видов происходят от предков, обитавших когда-то на травянистых формах.
Что касается горизонтального распространения, то в объеме всего пустынного комплекса чешуекрылых, оно показано в предыдущей главе. Здесь необходимо лишь кратко охарактеризовать с этой стороны каждую из экологических группировок. Из 26 псаммофильных видов 64% приходятся на среднеазиатских эндемиков, а ареалы большинства остальных, кроме Средней Азии, охватывают ближайшие к ней южные страны (Афганистан, Иран, Ирак). Только один вид доходит до Северной Африки, но ни одного нет в Южной Европе.
В группе эремофилов, несколько менее специализированных, чем псаммофилы, 35,9% от их общего числа приходится на эндемиков Средней Азии, 17,9% на формы, распространенные на территории Средней Азии, Афганистана, Ирана, Ирака. И только немногим более 1/4 видов этой группы (28%) имеет значительно более широкие ареалы, простирающиеся в пределах от Средней Азии до Северной Африки, Закавказья или также до Юго-Востока Европейской части Советского Союза. И совсем мало среди эремофилов форм, распространенных до южных областей Западной Европы.
В комплексе тугайных форм, занимающих в песках незначительное место ввиду немногочисленности видов и особей, один вид доходит до Закавказья, а два других до Южной Европы и Северной Африки.
Среди равнинно-горных ксерофилов нет эндемиков; ареалы более половины видов простираются и в Южной Европе. Формы, распространенные в пределах Средней Азии, Афганистана, Ирана и Ирака составляют 25% этого комплекса (относительно больше, чем в группе эремофилов, но в последней зато имеется значительное число эндемиков).
В группе эврибионтов, большинство которых для песков не характерны, также нет эндемичных форм. Ареалы всех видов обширные. Некоторые из них простираются до Южной Европы, но у остальных видов охватывают большую часть Палеарктики, или Голарктики, или даже еще шире.
При сопоставлении степени приспособленности чешуекрылых к жизни в песчаной пустыне (что в самых общих чертах отражено в приведенном выше разделении фауны на экологические группировки) с горизонтальным их распространением, становится очевидным, что чем более специализированы формы, т. е. чем более адаптированы к строго определенным условиям существования, тем ограниченнее их ареалы и, наоборот, наиболее лабильные виды, такие, как эврибионты, распространены особенно широко.
Однако здесь необходимо особо рассмотреть вопрос о вертикальном распределении форм, чтобы выяснить, какие же связи имеются между территориально соседними пустынной и горной фаунами.
Пределы вертикального распространения в Таджикистане видов, обитающих в песках Вахшской долины, представлены в табл. 3. Нижняя граница соответствует различным участкам обычно самой низменной части песков Юго-Западного Таджикистана; но в других местах Средней Азии эти же виды могут встречаться и ниже указанных высот. Приведенные в таблице данные основаны, главным образом, на сборах и наблюдениях автора, и высоты отражают не умозрительные обобщения, а фактические наблюдения. В табл. 4 показана верхняя граница распространения в Таджикистане видового состава экологических группировок песков низовья Вахша.
Таблица 3

Таблица 4
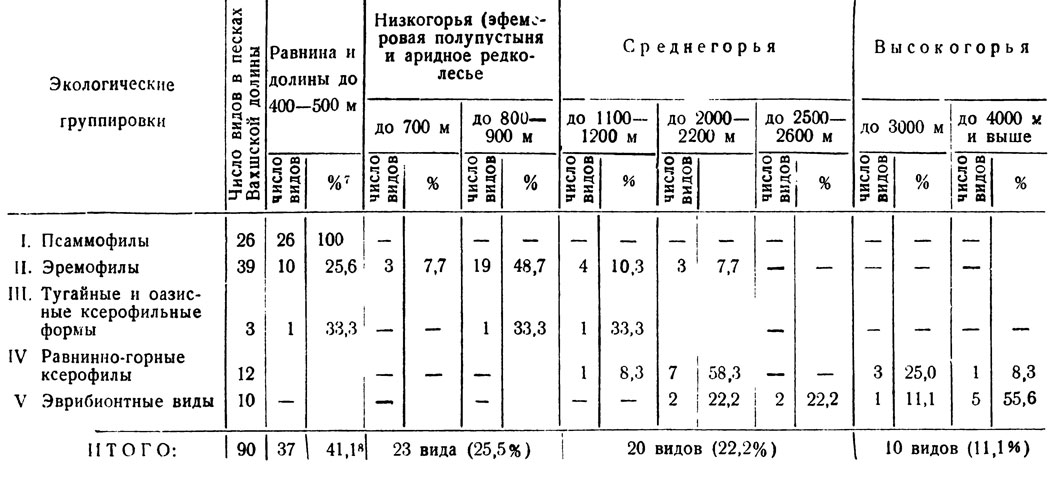
7(Проценты даны от общего числа видов данной экологической группы.)
8(В итоговой графе проценты даны от общего количества видов (90) в песках)
Из приведенных цифр видно, что очень большое количество форм приурочено исключительно к равнине и долинам, лежащим не выше 400 - 500 м над ур. м. В это число входят все псаммофильные виды, свыше четверти эремофилов и одна тугайная форма. Всего 37 видов, или более 41% от всех высших бабочек песков.
Более половины эремофилов (56,4% от общего объема этой группы) встречается в полосе сухих низкогорий с травянистой эфемеровой, аридной и субаридной древесно-кустарниковой растительностью, но не выше 700 - 900 м над ур. м. и у этого предела они обычно редки. В совокупности с долинно-равнинными формами этот низкогорно-равнинный комплекс составляет 66,7% (60 видов) от общего числа высших чешуекрылых, т. е. является самым обширным по числу видов и в основном состоит из наиболее характерных для песков форм.
В пределах среднегорий, но не выше 2600 м, отмечается 20 видов (22,2% всей фауны высших), причем псаммофилы там отсутствуют, и численно преобладают равнинно-горные ксерофилы (8 видов). Эремофилы количественно (7 видов) немного уступают им в этом поясе, но составляют лишь 18% всей группы эремофильного комплекса.
Наконец, в высокогорном поясе полностью отсутствуют как псаммофилы, так и эремофилы. Из двух представленных там группировок, общих с песками, - эврибионтов и равнинно-горных ксерофилов - количественно преобладают первые (6 видов), как наиболее лабильные и широко распространенные. Но всего в высокогорьях отмечается общих с песками форм лишь 11,1% (Ювидов).
В общем комплексе равнинных и равнинно-низкогорных форм, составляющем 2/3 высших бабочек песков, сосредоточены почти все среднеазиатские эндемики, а большинство остальных видов имеет более или менее ограниченные ареалы. Большинство форм, из 20, доходящих до среднегорий, отличается сравнительно широкими ареалами, захватывающими и Южную Европу; среди них только 2 эндемика (Cidaria tadzhikaria и Paraegle tesselata). Абсолютное большинство видов, достигающих высокогорий (до 3000 - 4500 м), распространено исключительно широко; но для песков эти формы не характерны.
На основании этих данных можно считать, что виды фауны песков тем выше поднимаются в горах, чем шире их ареалы и чем менее они специализированы. Это правило имеет некоторые исключения. Например, у Neolycaena tengstroemi и Satyrus enervate* ареалы сравнительно не очень велики (преимущественно среднеазиатские) но в горах эти виды доходят до 3000 м. С другой стороны, также в виде исключения, некоторые широко распространенные формы (Sideridis zeae, Melicleptria scutosa) в Средней Азии отмечены лишь в долинах не выше 700 - 800 м над ур. м. Однако в фауне бабочек песков нет узко локальных видов (таджикистанских или среднеазиатских эндемиков) с обширным диапазоном вертикального проникновения. Ни один псаммофил в условиях Таджикистана не проникает даже в низкогорьях (хотя не все псаммофильные виды связаны только с псаммофитными кормовыми растениями, и в некоторых горных районах встречаются небольшие участки песков), и нет ни одного характерного горного вида в песках. В среднегорном и высокогорном поясах (во всяком случае, выше 1200 м) совершенно отсутствуют представители таких политипических родов, как Armada, Anumeta, Thargelia, Anydrophila и др. столь характерных для жарких пустынь, особенно песчаных.
* (Но все-таки ареалы Neolycaena tengstroemi и Satyrus enervata никак нельзя назвать небольшими.)
Таким образом, анализ вертикального распространения обнаруживает, что у рассматриваемого песчаного фаунистического комплекса (фактически и у других биотопов Вахшской долины) очень мало общего с фауной среднегорий и, тем более, высокогорий. Если до среднегорного пояса небольшое число форм, более или менее локальных и характерных для пустынь, все же доходит, то почти все виды, общие и для высокогорий, являются широко распространенными и очень обыкновенными в различных биотопах. Слабость связей фауны зоны пустыни, среднегорий и высокогорий становится особенно заметной, если учесть чрезвычайную территориальную близость указанных поясов.
Из приведенных данных на материале по чешуекрылым становится очевидным тот факт, как это отмечается и по другим группам животного мира, что при зоогеографическом районировании необходимо подходить к фаунам гор и равнин дифференцированно, и проецирование фауны (или флоры) Средней Азии, и горной и равнинной, на единую плоскость лишь затемняет действительную картину распределения организмов. Рубеж, разделяющий горную фауну от равнинной в южных районах Средней Азии проходит в верхней части низкогорной полосы эфемеров и аридного редколесья, примерно на уровне 800 - 1000 м. В северных районах он, по-видимому, проходит по предгорьям. Этот рубеж, однако, является то более узкой, то более широкой полосой (с переходной фауной), высота которой над уровнем моря колеблется в зависимости от климатических условий местности.
Ввиду всего сказанного, для выяснения близких связей фауны песков Юго-Западного Таджикистана следует обратиться не к выше расположенным вертикальным поясам, а к пустыням Кара-Кум и Кзыл-Кум, лежащим примерно на тех же высотных уровнях, что и низовьях Вахша и Кафирнигана, хотя и удаленным от них на многие сотни километров.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://butterflylib.ru/ 'Чешуекрылые, бабочки, мотыльки, моли'