Чешуекрылые песков Вахшской долины - обедненный вариант фауны Кара-Кума. О принципах зоогеографического районирования
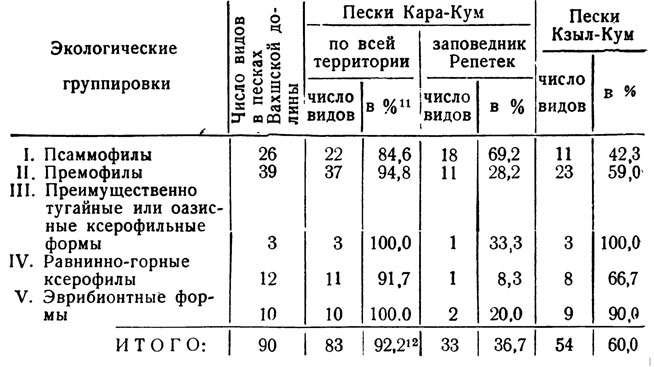
Даже беглого взгляда на список чешуекрылых вахшских песков достаточно, чтобы убедиться в большом сходстве этого комплекса с фауной Кара-Кума. Статистический анализ чешуекрылых Кашка-Кума Вахшской долины, Кара-Кума и Кзыл-Кума дан в табл. 5. Приведенные данные по Репетеку* - типичному участку песчаной пустыни - очень неполны, так как систематических сборов, охватывающих все сезоны, там не производилось. Сведения о фауне Кзыл-Кума еще менее полны, и для таблицы использованы, главным образом, довольно обстоятельные материалы, собранные на участках песков около очень удаленной от Южного Таджикистана восточной окраины Кзыл-Кума, а именно по Сыр-Дарье в районе станций Джулек, Байгакум и урочища Ай-Дарле (сборы В. Кожанчикова и др., хранящиеся в Зоологическом институте АН СССР).
* (Использованы материалы Зоологического института АН СССР, коллекция А. В. Цветаева и литературные данные.)
Таблица 5
10(Использованы материалы Зоологического института АН СССР, коллекция А.В. Цветаева и литературные данные).
11(Проценты даны от общего числа видов данной экологической группы.)
12(В итоговой графе проценты даны от общего числа видов (90) в песках Вахшской долины).
Подавляющее большинство видов (92,2%) фауны вахшских песков встречается и в Кара-Куме (но не всюду; многие виды - на окраине, около предгорий). В Кара-Куме отмечены почти все вахшские псаммофильные формы (22 вида, или 84,6%). Из их числа не найдено там пока 3 - 4 вида: Lemonia tancrei, Thargelia arenicola, Elaphria clara armeniaca и Armada kisilkumensis, но последний вид обычен в Кзыл-Куме и несомненно встречается хотя бы в Юго-Восточном Кура-Куме. Thargelia arenicola в той части Кара-Кума также, очевидно, имеется. Два других вида, Elaphria clara armeniaca и Lemonia tancrei. еще очень мало известны (первый из них до этой работы не был отмечен в Средней Азии, а второй - в Таджикистане). Очень вероятно, что они распространены в некоторых районах Кара-Кума и Кзыл-Кума. Так что фактически из вахшского комплекса в Кара-Куме, можно полагать, представлены все или почти все псаммофилы. Что касается эремофилов, то в Кара-Куме, особенно на южной окраине пустыни близ Копет-Дага, их относительно еще больше - 37 видов, или около 95% от общего числа форм этой группы. Там не найдены пока только Porphyrinia gratiosa Ev. и Cidaria tadzhikaria Stshetk. Остальные экологические группировки бабочек вахшских песков в Кара-Куме представлены почти в полном составе. Лишь из числа равнинно-горных ксерофилов в этой пустыне отсутствует Neoris stoliczkana, но она отмечена в Копет-Даге, и, возможно, местами заходит на окраину песчаной пустыни, где имеются насаждения кустарниковых видов Zygophyllum.
Если обратиться к очень неполным данным по Репетеку. расположенному в глубине песчаной пустыни Кара-Кум, то даже эти фрагментарные материалы свидетельствуют о большом сходстве видового состава бабочек Репетека, особенно псаммофилов, с фаунистическим комплексом вахшских песков. Замечательное сходство с Кашка-Кумом имеет и фауна весьма отдаленных от него районов Кзыл-Кума, где отмечено 54 общих вида (60%).
Возвращаясь к пустыни Кара-Кум, необходимо отметить, что, хотя ее фауна необычайно сходна по видовому составу с вахшскими песками, но она гораздо разнообразнее. В песках низовья Вахша среди чешуекрылых нет эндемичных для долины форм даже подвидового ранга (Thargelia arenicola Stshetkin, очевидно, встречается в Юго-Восточном Кара-Куме) и вместе с тем отсутствуют представители таких псаммофильных родов, характерных для Кара-Кума (или Кзыл-Кума) как Odontelia (O. margiana Pglr., O. megastigma Warr.), Borisia (B. hiemalis Fil.), Jaksarthia (J. elinguis Pglr., J. striolata Fill.), Marsipiophora (M. christophi Ersch.) и др.; выпадает ряд песчаных видов из псаммофильных или экологически более широких родов, например, Thargelia fissilis Chr.. Armada karakumensis Geras., A. secunda Ersch., Imitator ciliaria Men., Anydrophila simiola Pglr., A. mirifica Ersch., A. intermixta Warr., Rhodostrophia solitaria Chr., Lithostege excelsata Ersch. и др.* Из Cossidae нет Holcocerus pulverulentus Pglr. и многих других древоточцев, и т. д.
* (Marsipiophora christophi Ersch. и Armada karakumensis Geras. доходят в южном Узбекистане до низовья Сурхан-Дарьи (пески около г. Джар-Кургана), хотя там редки; но в Вахшской долине они отсутствуют.)
Таким образом, пески Вахшской долины представляют собой весьма заметно обедненный вариант Кара-Кума, обедненный главным образом, за счет ряда крайне специализированных форм, связанных с очень узким кругом кормовых псаммофитных растений. Однако, выше приведены данные по всему Кара-Куму, так как достаточно полных сведений по какому-либо их небольшому участку нет. В действительности пустыня Кара-Кум, простирающаяся на сотни километров очень неоднородна и включает как сыпучие, барханные, с крайне разреженной растительностью, так и в различной степени закрепленные грядовые и бугристые пески. Конечно, фауна любого сравнительно небольшого участка Кара-Кума менее разнообразна, чем пустыня в целом. Поэтому и фаунистическая обедненность вахшских песков относительно любого отдельного участка Кара-Кума оказывается фактически менее резкой, чем при сравнении с видовым составом всей пустыни.
Большое сходство фауны чешуекрылых вахшских песков с Кара-Кумом обусловлено тем, что Кашка-Кум в сущности является частью Кара-Кума, точнее - восточной оконечностью Юго-Восточного Кара-Кума. И нет ничего удивительного в том, что на далекой окраине Кара-Кума с обедененной растительностью и псаммофильная фауна оказывается не столь разнообразной, как в центральных районах. Даже в песках низовья Сурхан-Дарьи, лежащих недалеко от Вахшской долины, но ближе к Кара-Куму, встречается ряд видов, отсутствующих в Кашка-Куме.
Итак, пески Кашка-Кум, их растительность и фауна являются частью обширной пустыни Кара-Кум и ее северо-восточного продолжения - Кзыл-Кум. Но если это бесспорно, то в таком случае возникает вопрос: есть ли серьезные основания "разрывать" на части пустыню Кара-Кум, как это делают при флористическом районировании некоторые ботаники? Выше было показано, что связь фауны песков с высокогорным или среднегорным миром чешуекрылых очень слабая, хотя пространственно они соседствуют друг с другом. Однако ботаниками объединены в Южнотаджикистанском флористическом районе, в сущности, различные флоры. Здесь - и псаммофитная растительность песков низовий Вахша и Кафирнигана, и низкотравная эфемеровая пустыня (низкотравная полусаванна), и низкогорья с аридным редколесьем (шибляк), и среднегорья с фрагментами мезофитной лесной растительности и даже арчовников. Таким образом, в одном каком-либо флористическом подрайоне объединены различные вертикальные пояса от равнин и низких долин с высотами 300 - 400 м над ур. м. до среднегорий или до высокогорий включительно. Вахшско-Кафирниганский и Пархаро-Кировобадский подрайоны, например, отличаются только тем, что в первом есть пески с псаммофитной растительностью (Кашка-Кум), а во втором они отсутствуют. При этом вахшские и кафирниганские пески "отрываются" от Кара-Кума и искусственно объединяются с горным ландшафтом, в котором высоты достигают 2500 м. Конечно, могут "резонно" возразить, что все дело в другом подходе к районированию, дело, мол, в различных принципах. Однако, на мой взгляд, при биорайонировании нельзя придерживаться одних и тех же принципов, как в случае выделения обширных территорий земной поверхности (например, таких зоогеографических областей, как Палеарктика и т. д.), так и при дроблении на мелкие единицы - районы и подрайоны. Если при вычленении обширных зоогеографических или флористических территорий земной поверхности метод районирования только по типам ареала без учета экологической дифференцировки флоры и фауны оправдывает себя, то строгое следование этому принципу при более мелком дроблении приводит к явному противоречию с фактами*. В этом случае, как это было на примере вахшских песков, единая флора (или фауна) расчленяется, а мало родственные флористические (фаунистические) комплексы различных высотных поясов, наоборот, объединяются. Современные ареалы животных и растений обусловливаются не только историей организмов, но, в первую очередь, экологическими факторами. Большинство ботаников и зоологов не отрицают значения этих факторов в распространении видов, но многие при районировании на деле игнорируют это беспорносположение. Одним из наиболее ярких и последовательных представителей этого течения в биогеографии, до недавнего времени господствовавшего, был известный ботаник Е. В. Вульф. Вот что он писал, излагая свою точку зрения: "Отделить флоры вершин от горных склонов не представляется возможным, так как одна переходит в другую, представляя собой результат естественного видоизменения флоры под влиянием смены экологических условий по мере подъема от уровня моря к вершине хребта. Эта дифференциация флоры горных возвышенностей на пояса имеет место сейчас даже в тропиках и существовала, конечно, и во все геологические периоды, в том числе и в третичном периоде, о чем свидетельствуют палеоботанические данные, как мы это сейчас увидим.
* (При выделении обширных областей по типам ареалов фактически учитываются как генезис фауны, так и экологическая обстановка, ибо большие территории (например Палеарктическая область) имеют и свои экологические особенности, резко отличные от таковых других областей.)
И если часть этих горных флор и не представляет собой видоизменения средиземноморских типов, а мигрировала сюда по горным хребтам или проникла с севера во время ледниковых периодов, то это еще не дает основания для отнесения их к тем областям, откуда эти виды мигрировали.
Таким образом, с нашей точки зрения, ни экологический тип растительности, ни характер климатических условий не могут быть решающими для установления границ флористических (но не растительных) областей. Таким критерием должна быть история этой флоры, выявляющаяся в типе ареалов ее видов" (Вульф, 1944, стр. 133 - 134).
Итак, флора, ее история, отражающаяся на ареалах видов, рассматриваются Вульфом вне климатических и экологических условий развития. И хотя автор не отрицает дифференциации флоры горных областей на пояса как в настоящем, так и в геологическом прошлом земли, однако он, исходя из "исторической концепции", рассматривает обширную территорию древнего Средиземноморья, включая и всю Среднюю Азию, как единую, не расслаивая ее по вертикали, и подразделяет флору, в сущности, чисто географически. Характерно, что в Средиземноморскую область вошла не вся территория древнего Тетиса (не вошла даже Палеарктическая часть умеренной зоны Европы), так как Вульф вслед за М. Г Поповым и другими его предшественниками считает важнейшим моментом для установления границ флористических областей тип ареалов. Однако, выясняя типы ареалов, автор рассматривает флору не каждого пояса в отдельности, а всю в целом, смешивая ее в единый конгломерат. Понятно, что при таком подходе к проблеме результаты будут иными, чем при дифференцированном анализе. А между тем, если наблюдается переход флоры одной в другую нижнего и вышележащих поясов, на чем делает ударение Е. В. Вульф, то следует также отметить, что внутри каждого вертикального пояса по его горизонтальному простиранию также имеются переходы, и они несравненно более плавные, чем между соседними поясами (см. табл. 4 и 5). Фауна чешуекрылых восточной окраины Кзыл-Кума имеет большое сходство во всех отношениях с фауной вахшских песков (общие виды составляют 60%), лежащих за тысячу с лишним километров, тогда как между фаунами песков Вахшской долины и ближайщих среднегорий общего чрезвычайно мало (общие виды составляют 22,2%, причем многие из них в среднегорьях редки и для них не характерны. В песках Кашка-Кум имеется некоторое число видов общих и для пустынных районов Северной Африки или Монголии, еще больше эти фауны роднят многие близкие пустынные виды и роды. В соседних же с Вахшской долиной высокогорных районах нет ни одного представителя псаммофильной или вообще пустынной фауны. И, конечно, в высокогорной фауне Зеравшанского или Гиссарского хребтов имеется гораздо больше общих (и к тому же характерных) элементов с верхним поясом Северо-Западных. Гималаев, чем с ближайщей окраиной Кзыл-Кума или эфемеровой подгорной пустыней. Е. В. Вульф признает существование вертикальной поясности в прошлые геологические эпохи, но не делает из этого вывода о большом значении вертикальной дифференциации флоры в современном распространении видов. Из его слов вытекает, хотя он этого прямо и не говорит, что процесс видообразования (и изменение в результате этого фауны) происходит с большей скоростью, чем расселение организмов в пространстве по экологически подходящим нишам. А между тем в современную эпоху (и нет оснований утверждать, что было иначе в прошлом) видообразование в целом происходит, несомненно, в более замедленном темпе, чем расселение организмов и изменения их ареалов. Сторонники указанной "исторической концепции" подходят к районированию чисто умозрительно, формалистично, поскольку они не считаются с экологической обстановкой, в которой протекает развитие вида и флоры в целом. Есть два обстоятельства, способствующие распространению этого течения в биогеографии. Во-первых, чаще всего полного анализа флоры не производится и читателю преподносится готовая схема районирования, подкрепленная очень немногими примерами. Это и понятно: представить флору в цифровом выражении, в соотношениях различных элементов, дело крайне трудоемкое, поскольку касается многих сотен или тысяч видов; к тому же часто отсутствует более или менее полный список не только растительности каждого биотопа, но и флоры данной территории в целом, и авторы вынуждены ограничиваться лишь некоторыми примерами. Во-вторых, в горных местностях действительно часто (но далеко не всегда) имеет место совпадение внутрипоясных границ на разных высотах, в результате чего намечается как бы общая вертикальная граница для нескольких или даже всех поясов, от вершины до равнины или долины. Это вполне понятно, так как поверхность суши, будь то равнина или, тем более, горный ландшафт, не является однородной и расчленена на отдельные более или менее географически обособленные участки. В таких до известной степени обособленных местностях наблюдаются свои особенности (в большей или меньшей степени) климата, которые нередко проявляются не в одном, а в ряде вертикальных поясов. Это всем известное явление наблюдается как на небольших площадях, так и на обширных участках земной поверхности. Естественно, климатические особенности каждой местности отражаются на составе ее флоры и фауны. Сторонники исторического метода и производят чаще всего биорайонирование по этим географо-климатическим границам. И так как детального статистического и генетического анализа всей флоры (или фауны) с приведением конкретных цифровых соотношений и с учетом экологического и вертикального распределения организмов обычно не делают, а ограничиваются лишь отдельными примерами, которые нетрудно подобрать, то упускают из вида или не придают значения тому обстоятельству, что поясные различия в составе флоры и фауны той или иной местности, как правило, во всех отношениях несравненно более резкие, чем изменения в горизонтальном направлении. В результате этого районирование проводится не путем поясного расслоения горных территорий и последующего выделения более мелких флористических (или фаунистических) единиц внутри пояса, а методом вертикального деления территорий на отдельные участки, включающие различные вертикальные пояса от жарких долин до снежных высокогорий. Могут возразить, что одни принципы у ботаников, другие у зоологов. Однако, вертикальное распределение организмов, будь то животные или растения, подчинено одним и тем же закономерностям и, хотя в каждой группе имеются свои особенности, они не столь велики, чтобы исследователи пришли к диаметрально противоположным выводам. Подтверждением сказанному является и то, что сторонники экологического и "исторического" подхода имеются как среди ботаников, так и среди зоологов. Чисто "исторический" подход к проблеме биорайонирования, как это имеет место у Вульфа, односторонен и дает искаженное представление о распределении организмов по земной поверхности; экологический же принцип районирования отражает не только современное распространение видов, но фактически и генезис фауны или флоры. При зоогеографическом анализе генезис фауны не может быть исключен, но когда при рассмотрении современного распределения организмов руководствуются единственно "историческим" фактором (и к тому же в таком аспекте, как у Вульфа), или считают его главным, то это приводит к поразительным искажениям.
Всякое районирование земной поверхности является, конечно, целевым. Зоогеографическое деление суши должно отражать в первую очередь действительную картину распределения животных организмов по земной поверхности; оно должно основываться на детальнейшем изучении горизонтального и вертикального распределения и экологической приуроченности организмов. Однако, это важнейшее условие, как было сказано, нередко до сих пор многими биогеографами нарушается, особенно в применении к горным странам, а районирование в таком случае становится чисто географическим и теряет свой смысл; истинный характер распределения организмов остается не вскрытым.
Главная ошибка многих ботаников и зоологов состоит, пожалуй, даже не в том, что они подходят к решению проблемы с позиций "исторической концепции" (ибо экологический метод не отвергает, а наоборот, вскрывает генезис фауны), а в том, как эта концепция у них преломляется, в том, что они априорно считают формирование фауны горных гребней, склонов и подножных равнин как процесс переработки единой равнинной, и только равнинной, фауны, происходящий с момента поднятий горных сооружений; они полагают, что проникновение других фаунистических элементов из соседних стран и из уже существующих гор - явление второстепенное, не имеющее большого значения в формировании фауны того или иного пояса, т. е. рассматривают процесс возникновения новых видов, скажем горных из долинных, как протекающий более ускоренно в сравнении с расселением и проникновением видов по соответствующим их природе экологическим путям.
Именно такой точки зрения и придерживался из зоологов П. А. Семенов-Тянь-Шанский (1937). Собственно, эта концепция в той или иной форме была общепринятой до самого последнего времени. Из лепедоптерологов, касавшихся районирования Средней Азии, ее придерживались Н. Я. Кузнецов (1929) и И. В. Кожанчиков (1937). И если в настоящее время большинство зоологов при зоогеографическом расчленении обширной равнинной части СССР подходят к проблемме в общем правильно, поскольку в этом случае учитывается ландшафтная зональность, то при районировании горных областей, таких, как Средняя Азия, вертикальная поясность обычно фактически игнорируется. Правда, многие зоологи сейчас, отмечая коренные различия низинной и горной фаун, выделяют их или в отдельные провинции одной подобласти или в разные подобласти (Кузнецов, 1950; Крыжановский, 1953: Борхсениус, 1957; Нарзикулов, 1962 и др.), однако вычленения вертикальных поясов в пределах Средней Азии, именно как зоогеографических единиц, до сих пор не сделано и в сущности эта проблема в энтомологической литературе не обсуждалась.
Между тем еще в 20-е годы появились первые указания на сходство высокогорной фауны (птиц) различных горных сооружений Палеарктики. В частности, в 1922 г. Штреземан (Stresemann) установил, что фауна птиц альпийского пояса Европы резко отличается от равнинной орнитофауны и вместе с тем имеет тесную связь с фауной нагорья Центральной Азии. П. П. Сушкин в 1928 г. привел неоспоримые доказательства единства фауны птиц всех высокогорий Северного полушария.*
* (Эти данные Штреземана и П. П. Сушкина приводятся по Штегману (1937))
Наконец, Б. К. Штегман (1937) подчеркивает различный характер орнитофауны высокогорных областей Палеарктики и нижележащих поясов. Он чрезвычайно близко подошел к пониманию необходимости поясного расчленения фауны. И все же окончательно эта мысль у него не оформилась. На его карте (Штегман, 1938) типы орнитофауны Палеарктики показаны казалось бы очень наглядно, но слишком схематично, и его метод нанесения на карту типов фаун не позволил разобраться с такой сложной территорией, как Средняя Азия, хотя все же высокогорную фауну он выделил. В представлении Штегмана, орнитофауна Средней Азии смешанная и только высокогорная явно обнаруживает тесные связи с тибетской фауной.* Штегман (1937) подчеркивает необходимость глубокого изучения закономерностей распространения организмов и убедительно доказывает, что нельзя статистическим методом анализировать фауну всей обширной местности, так как в общем фаунистическом списке смешиваются представители различных фаун, в результате чего картина распространения животных лишь затемняется. Однако он не довел эту идею до конца и вообще отказался от районирования, хотя мысль о внутрипоясном зоогеографическом районировании казалась бы сама напрашивается из его же доказательств. Ведь дело не в статистическом методе. Этим методом не следует пренебрегать. Он дает конкретный цифровой материал для выявления сходства и различия фаун. Дело не в статистике, а в том, как ею пользоваться. Если анализировать фауну, включающую различные пояса в целом, то статистика не поможет выяснить закономерности распределения организмов. Но, если таким методом проанализировать фауну каждого вертикального пояса в отдельности, а в ряде случаев это необходимо проделать и в отношении отдельных биотопов, то результат будет совсем иной, и в этом случае картина распределения организмов станет предельно ясной.
* (Для нас важно у Штегмана не то, что по его представлениям, высокогорная фауна Средней Азии тяготеет к Тибету (в этом отношении большинство энтомологов, по-видимому, с ним расходятся), а понимание им общности происхождения вообще всей высокогорной фауны и глубоких отличий ее от фауны нижележащих поясов.)
Из лепидоптирологов, расчленяющих фауну по вертикальным поясам, т. е. подошедших к решению проблемы зоогеографического районирования наиболее близко, необходимо отметить А. И. Куренцова. Но его работы касаются Дальнего Востока и не затрагивают Средней Азии.
В задачу данной работы не входит зоогеографическое районирование Таджикистана или более обширных областей. Однако необходимо кратко отметить, куда же следует отнести фауну Кара-Кума, в которую нужно, как выше было установлено, включить и вахшский комплекс чешуекрылых? Б. А. Кузнецов (1950) на материале по млекопитающим относит пустыню Кара-Кум к подобласти степеней и пустынь Палеарктики. Материал по чешуекрылым не дает оснований для объединения степей с пустынями в единую подобласть; но та часть подобласти, которую у Б. А. Кузнецова занимают пустыни, очерчена им детально и в соответствии не только с распространением млекопитающих, но и беспозвоночных. Во всяком случае, в таких границах подобласть пустынь приемлема при зоогеографическом районировании на основании распространения чешуекрылых. Что касается гор Средней и Центральной Азии, то они, с моей точки зрения, очевидно, должны быть расчленены по вертикальным поясам, а масштабы границ между ними определятся анализом фауны.* При таком расслоении фауны по вертикальным поясам необходимо учитывать весь видовой состав, а не отдельные формы - индикаторы.
* (Из предварительного подсчета видов дневных бабочек (Rhopalocera) на различных высотах Гиссарского хребта оказалось, что в высокогорном поясе (от 3300 м и выше) около 70% всего видового состава составляют формы, отсутствующие на более низких высотах. В среднегорной полосе от 1700 до 2500 м подобных "поясных эндемиков", отсутствующих в высокогорьях и низкогорьях, насчитывается свыше 54%. Так что даже самые предварительные подсчеты уже подтверждают изложенные выше соображения о значительной обособленности фаун каждого пояса.)
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://butterflylib.ru/ 'Чешуекрылые, бабочки, мотыльки, моли'