Типы годичных циклов развития и распределение чешуекрылых по сезонам года
При систематической, ежедневной, ловле бабочек в жарких долинах Средней Азии у наблюдателя часто создается впечатление, что в течение вегетационного сезона, с ранней весны до глубокой осени, видовой состав непрерывно изменяется. Почти каждую неделю что-нибудь добавляется новое к сборам, что-то исчезает. Однако анализ динамики видового состава убеждает в том, что в действительности фауна по сезонам года распределяется очень неравномерно и что в определенные периоды она относительно статична, тогда как в другие сезоны, наоборот, количество видов резко возрастает или убывает. В фауне песков Вахшской долины моновольтинные виды значительно преобладают над поливольтинными и составляют 66,7% (60 видов), последних же насчитывается лишь 30 видов, или 33,3 % от общего числа форм высших чешуекрылых. Рассмотрим сначала группу поливольтинных видов. В жестких условиях существования, в песках, где лето отличается очень высокими дневными температурами и большой сухостью воздуха, полицикличные формы могут развиваться только на кустарниках и многолетниках. Если некоторые и встречаются на эфемеровых растениях, то только в период роста и цветения последних - весной и в начале лета; развитие последующих поколений этих форм, начиная с третьего, происходит вне этого биотопа, в поливной полосе или в тугайной пойме. Такими временными обитателями песчаной пустыни являются широко распространенные полифаги - Pieris rapae debilis, Colias erate, Pyrameis cardui и, очевидно, Scotogramma trifoli. Быть может, к этой категории относится и ряд других видов, в частности Papilio machaon centralis, отмеченный в песках в первых двух генерациях. Но вероятнее всего этот вид в фазе куколки имеет летнюю (после 2-го поколения) диапаузу. Гусеницы второй генерации Pyrameis cardui в песках до конца мая питаются однолетними видами Cousinia (из сем. Compositae) и Arnebia baldshuanica Lipsky (сем. Borraginacea); Colias erate может развиваться на травянистых астрагалах или других однолетних бобовых, Pieris rapae - на различных крестоцветных, Papilio machaon - на видах Haplophyllum (из Rutacea) и, вероятно, на некоторых зонтичных и сложноцветных. Эти виды в поливной полосе долины (кроме P. machaon, который там не найден), где в течение всего летнего сезона имеются вегетирующие растения, дают до 4-6 генераций. Однако большинство поливольтинных видов является постоянными обитателями песков. Среди поливольтинных имеются формы, более или менее широко распространенные и свойственные различным биотопам, в том числе тугаям и полосе орошаемых земель. Значительное число однако характерно лишь для пустынного ландшафта, в частности для песчаной пустыни.
Таблица 8
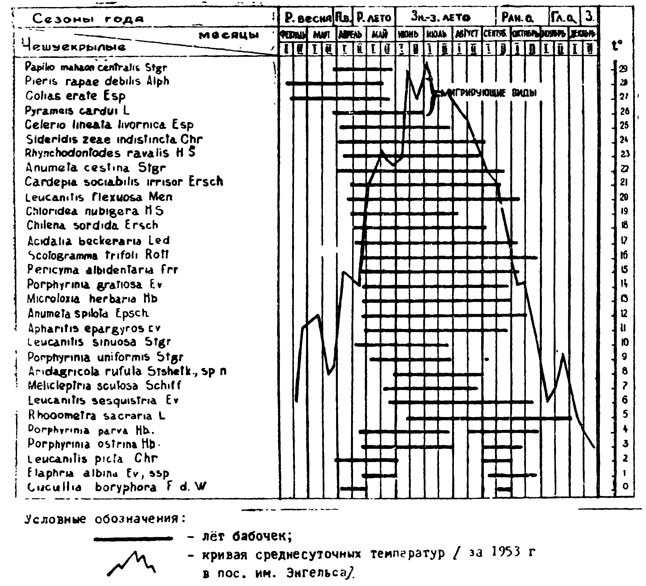
По характеру годичного цикла развития поливольтинные чешуекрылые в песках представлены двумя типами: 1) с непрерывным рядом поколений в течение теплого времени года и 2) с рядом поколений, прерывающимся в период знойно-засушливого лета диапаузой в фазе куколки. Формы с непрерывным рядом поколений составляют подавляющее большинство. В течение теплого времени года они развиваются в трех - четырех генерациях, число которых обусловлено генетическими особенностями вида и отчасти погодными условиями года. Зимовка большинства видов происходит в фазе куколки (у некоторых зимуют гусеницы, например, у Anumeta spilota); лёт большей части форм первого поколения начинается сравнительно поздно: в первой половине апреля или в конце этого месяца, а у некоторых (Leucanitis sesquistria и др.) даже в конце мая (см. табл. 8 - 9 и 13). Это относится к формам с обоими типами цикла развития, поэтому в табл. 9 все поливольтинные формы объединены вместе. Лёт последней генерации заканчивается у большей части видов во второй половине сентября или в октябре, реже - в июле, августе - начале сентября (табл. 9). Только один вид (Rhodometra sacraria), характерный для поливной полосы и речной поймы, продолжает летать глубокой осенью до начала декабря.
Таким образом, у большинства поливольтинных форм лёт бабочек и откладка яиц начинается с конца весны или в начале раннего лета и продолжается до первой половины ранней осени, т. е. приходится на период с момента распускания листвы и достаточного развития побегов кустарниковой псаммофитной растительности и травянистых многолетников, когда развивающиеся на них виды могут быть обеспечены кормом (табл. 9). Начало лёта бабочек первой генерации Anumeta cestina совпадает с появлением самых первых зеленых веточек кормового растения Calligonum griseum; другой близкий вид, Anumeta spilota - потребитель того же джузгуна, появляется несколько позже, когда кустарник достаточно хорошо распустится. У ряда видов (Apharitis epargyros, Chilena sordida, Pericyma albidentaria, Leucanitis flexuosa и Rhynchodontodes ravalis), питающихся верблюжьей колючкой (Alhagi canescens и близкими видами), лёт наступает еще до цветения кормового растения, но когда вегетационные органы достаточно хорошо разовьются.
Полицикличные формы в имагинальной фазе составляют, главным образом, летний аспект фауны песчаной пустыни. Заметное число их имеется также в поздне-весенней фауне и в начале ранне-осеннего периода. Но ранней весной (вторая половина февраля, большая часть марта) и во второй половине ранней осени (вторая половина октября - первая половина ноября) поливольтинных представителей мало, а глубокой осенью встречается только один вид. Если подсчитать число летающих поливольтинных видов по месяцам, то оказывается, что наибольшее видовое разнообразие (23 - 28 видов) наблюдается в период со второй половины апреля, т. е. момента наступления раннего лета, до конца июля (до середины знойно-засушливого сезона), когда суточные температуры, достигнув максимума, начинают понемногу снижаться. С этого момента из комплекса полицикличных форм медленно, но неуклонно начинают выпадать отдельные виды (табл. 8 и 13). Этот, процесс резко ускоряется с конца сентября, так что к концу октября поливольтинные формы исчезают почти полностью (кроме Rhodometra sacraria).
Таблица 9
Поливольтинные виды с другим типом цикла составляют очень небольшую группу. У этих форм в фазе куколки наблюдается летняя диапауза, а в лёте бабочек вследствие этого - резкий разрыв в жаркое время года, в июле - августе (El. albina, L. picta) или с начала мая до третьей декады сентября (Cucullia boryphora). К указанной группе, кроме Cucullia boryphora, Elaphria albina и Leucanitis picta, принадлежит, быть может, Porphyrinia parva и, вероятно, Porphyrinia ostrina. У последней, если у нее действительно имеется летняя диапауза*, этот разрыв значительно меньше и приходится на август, а у P. parva - не более 20 дней (в конце июля - первой половине августа). Leucanitis picta развивается в двух поколениях. Часть куколок второй генерации дает бабочек уже в начале июня, но большинство их вылупляется (после диапаузы куколок) с начала сентября до середины октября. Наличие летней куколочной диапаузы можно определить и без воспитания гусениц, по характеру лёта бабочек. Биология и преимагинальные фазы Cucullia boryphora остаются неизвестными. Бабочки летят дважды, и это наблюдается в различных местностях и в разные годы: в апреле и после длительного перерыва во второй половине сентября. При этом сентябрьские особи всегда совершенно свежие и, следовательно, они не могли появиться после имагинальной диапаузы, как это наблюдается у моновольтинной Euxoa conspicua и некоторых других видов.
* (Во всяком случае, в августе P. ostrina не встречалась в долине. Даже, если она в это время просто очень редка, то, очевидно, это свидетельствует о том, что часть куколок в августе впадает в диапаузу.)
Летняя диапауза куколки, как и у моновольтинных форм, является приспособлением к переживанию неблагоприятных для вида крайне высоких летних температур и большой сухости воздуха. Кормовые растения здесь, по-видимому, роли не играют, так как Leucanitis picta, например, обеспечена кормом в течение всего летнего периода (на сером джузгуне развиваются и другие виды совок, но они не имеют летней диапаузы) и тем не менее, у нее, с началом знойно-засушливого лета наступает диапауза, приуроченная именно к этому моменту.
Возникает вопрос: какие факторы вызывают летнюю диапаузу? Специальных исследований в этом направлении по среднеазиатским видам не проводилось. Однако можно полагать, что в приведенных примерах диапауза вызвана непосредственно летними высокими температурами. Характерно, что у L. picta она наступает, когда среднесуточные температуры поднимаются выше 22 - 23°С (в начале июня) и снижается в начале сентября, когда температуры вновь спускаются примерно до того же уровня, до 22 - 23°С. В лабораторных условиях, где гусеницы содержались при более низкой температуре воздуха, диапауза наступала позже (лишь у особей, окуклившихся после второй декады июня), хотя длина дня в помещении была такой же, как и в природе. Однако не исключено, что диапауза упомянутых видов обусловлена укорочением дня. Дело в том, что у некоторых видов гусеницы, ведущие обычно ночной образ жизни, весной и даже в начале раннего лета, а также осенью, в первой половине дня, а то и полные сутки, могут оставаться на кормовых растениях (например, у той же L. picta), но с наступлением жары они все рано утром покидают растения и закапываются глубоко в песок. Длина дня для них таким образом сокращается. Окончательно этот вопрос можно решить лишь экспериментальным путем. Но скорее всего, верным окажется первое предположение, так как число гусениц, остающихся днем на кормовых растениях, невелико, и, очевидно, в это время суток представлена не вся популяция. Об этом же свидетельствуют и результаты воспитания в лаборатории, приведенные выше.
Что касается осенне-зимней диапаузы полицикличных форм, то А. С. Данилевский (1961) убедительно показал, что у многих чешуекрылых она возникает при сезонных изменениях длины дня до определенной пороговой величины. Большинство поливольтинных видов песчаной пустыни, очевидно, имеет длиннодневный тип развития, когда диапауза наступает в осеннее время при уменьшении долготы дня до пороговой для данного вида величины. Этот момент у основной массы видов рассматриваемого комплекса, приходится на период с сентября до конца октября. Следует иметь в виду, что вопрос о фотопериодических реакциях у многих пустынных видов осложняется биологическими особенностями их личиночной фазы. Имеются в виду виды, у которых гусеницы ведут ночной образ жизни, а днем скрываются, закопавшись глубоко в песок. У некоторых форм, например у Anumeta spilota, молодые гусеницы ведут дневной образ жизни, взрослые же ночной.
Если у подавляющего большинства рассмотренных полицикличных форм зимовка происходит чаще всего в фазе куколки, а развитие гусениц и лёт бабочек в непрерывном ряде поколений приходятся на период с поздней весны до ранней осени, захватывая весь или почти весь сезон знойно-засушливого лета, то распределение активных и покоящихся фаз по сезонам года у представителей вдвое более обширной группы моновольтинных форм оказывается совсем иным. Среди моноцикличных чешуекрылых очень мало широко распространенных видов; во всяком случае, процент их по сравнению с таковым в комплексе поливольтинных форм не велик. Здесь больше эремофильных форм (в абсолютном и процентном выражении) и сосредоточена основная масса типичных псаммофилов. Большинство видов питается травянистыми однолетними или эфемероидными растениями. Общим для всех форм оказывается то, что фаза гусеницы приходится на время бурного роста и развития растительности - весной и у части видов до начала раннелетнего сезона. В период знойно-засушливого лета все виды находятся в состоянии диапаузы на различных фазах развития, специфичных для данного вида. У большинства летняя жара и зимние холода переживаются в фазе куколки, реже в состоянии яйца. Характерной особенностью в лёте моноцикличных форм является также приуроченность их лёта или к весенне-ранне-летнему, или к осеннему сезонам, и полное отсутствие бабочек во второй половине знойно-засушливого лета,* если не считать, конечно, небольшого числа диапаузирующих в имагинальной фазе видов (табл. 10 и 13). Этот "вакуум" жаркого периода заполняется, как было показано выше, поливольтинными формами.
* (При разграничении сезонов года использованы терминология и схема деления, предложенная Г Э. Щульцем (1950) для Гиссарской долины. Мной внесены поправки и дополнения в соответствии с климатическими условиями Вахшской долины и фенологией чешуекрылых.)
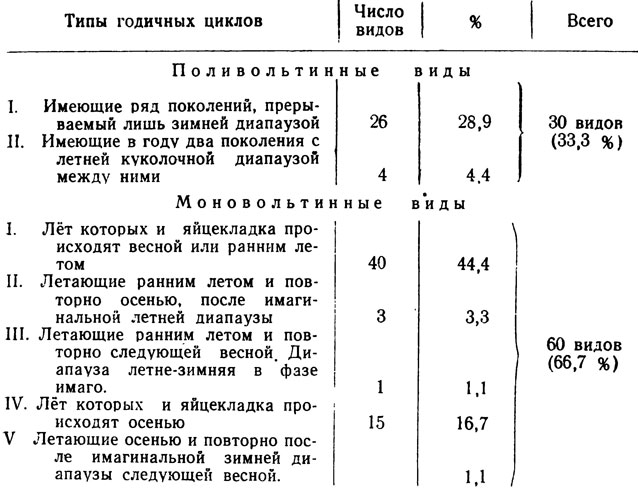
Моноцикличные виды по характеру лёта и типу цикла развития распадаются на 5 количественно неравных групп: 1) приуроченные к весенне-ранне-летнему периоду, 2) летающие дважды - ранним летом (после выплода) и осенью после летней имагинальной диапаузы, 3) летающие дважды - ранним летом и вторично следующей весной после имагинальной летовки и зимовки, 4) приуроченные к осени, 5) летающие осенью и второй раз весной после имагинальной зимовки (табл. 10).
Первая группа форм, летающих в весенне-раннелетний период, по числу видов (40) самая обширная. Личиночная фаза приурочена ко времени роста и цветения весенних травянистых растений (эфемеров и эфемероидов), которыми, главным образом, и питаются гусеницы. Летняя диапауза и зимовка у большинства видов происходит в фазе куколки или, у некоторых, в состоянии яйца. В Вахшской долине в апреле дожди уже не часты и для гусениц не страшны, но травянистая растительность в результате предшествовавших февральско-мартовских более или менее обильных осадков бурно развивается. В начале мая она выгорает, но к этому времени даже у наиболее поздних видов заканчивается развитие и происходит окукление. В этой фазе пустынные виды могут пережить самое неблагоприятное время года - летний зной и зимние холода. Поэтому эти формы вылетают из куколки сравнительно рано весной, как раз в период наилучшего развития их кормовых растений. Zegris fausti связана с крестоцветным Crambe Kotcshyana Boiss.; вылетая в конце марта, она уже находит для откладки яиц розетки листьев этого растения; Euchloe belia, живущая на эфемеровых крестоцветных появляется еще раньше; Cucullia chamomillae летает в период развития и цветения основного там ее кормового растения - мелкой ромашки (Matricaria lamellata Bge.) в апреле.
Таблица 10

Некоторые виды весеннего комплекса (например, Oria musculosa) перезимовывает в фазе яйца. У таких форм имеется потенциальная возможность к еще более раннему весеннему отрождению гусениц. Однако эта возможность фактически реализуется в соответствии с сезонным состоянием кормовых растений. Выплод гусениц, их окукление и дальнейшее развитие синхронно связано с сезонными изменениями в природе. У Oria musculosa гусеницы с ранней весны обеспечены кормом, но питаются преимущественно более или менее крупными злаками (особенно культурными), имеющими достаточно толстые стебли, внутри которых они живут. Ввиду того, что питание и рост гусеницы приходится в основном на период, когда стебли более или менее сформированы, а развитие куколки (до 25 суток и даже более) довольно долгое, то и лёт бабочек в Вахшской долине начинается сравнительно поздно - с середины мая до конца месяца. Впрочем, Oria musculosa в песках редка и встречается только на их окраине, так как подходящих кормовых растении там очень мала.
В составе рассматриваемой группы очень мало широко распространенных форм, преобладают пустынные эндемичные и относительно локальные с ареалами не выходяoими за пределы Средней Азии, Афганистана к Ирана; среднеазиатских эндемиков 24, т. е. 60% всей группы. Это не удивительно, так как распространение пустынных видов определяется в значительной степени характером пищевых связей (все или подавляющее большинство форм - монофаги или олигофаги) и адаптацией их к жестким летним климатическим условиям существования пустыни и соответствующему сезонному ритму [35% составляют псаммофилы (14 видов), 56,6% приходится на долю эремофилов (22 вида)].
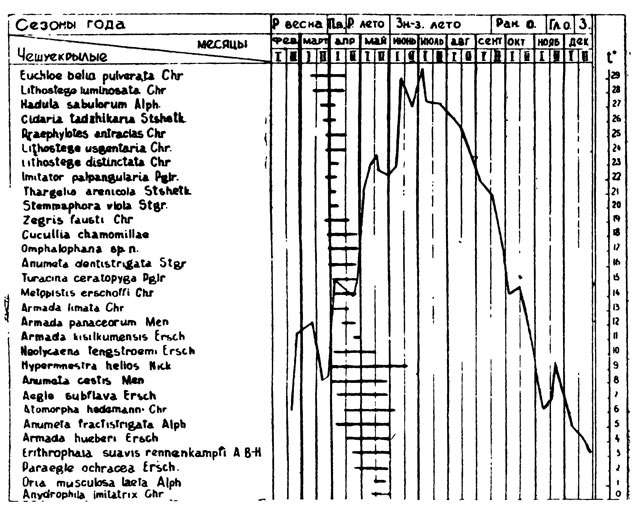
Как было сказано, лёт видов с данным типом годичного цикла происходит весной или в период раннего лета. На табл. 10 отчетливо видна приуроченность лёта значительной части видов к концу марта - первой половине апреля, т. е. к периоду поздней весны, когда происходит массовое цветение эфемеровой и эфемероидной растительности. Другой большой ряд видов приходится преимущественно на вторую половину ранне-летнего сезона (в основном, во второй половине мая), когда среднесуточные температуры резко повышаются, а весенняя травянистая растительность выгорает и отмирает или переходит в фазу покоя (эфемероиды). Многие формы однако летают в течение всего апреля, или всего мая, или на протяжении значительной части того и другого месяца. У некоторых лёт длится с апреля или с мая почти до конца июня. При этом происходит захождение лёта различных видов, и наибольшее число весенних видов отмечается в апреле и мае, когда количество всех моновольтинных форм (включая представителей и из других групп) достигает соответственно 33 и 25, или в любые полмесяца от 23 до 25 видов (табл. 10). Из дневных бабочек весной летают Euchloe belia, Zegris fausti, Praephylotes antracias (и кроме того, несколько обычных поливольтинных видов - Papilio machaon, Pieris rapae, Colias erate и Pyrameis cardui). Из ночных видов очень характерны для этого времени Thargelia arenicola, Imitator palpangularia, Omphalophana sp., Turanica ceratopyga, Anumeta dentistrigata, Armada limata, Arm. kisilkumensis, Metopistis erschoffi, ряд пядениц - Lithostege luminosata, L. usgentaria, L. distinctata и многие другие виды (табл. 10). Для раннего лета (май) характерны Paraegle ochracea, Metopoplus alboflavicola, Anydrophila imitatrix, Epharmottomena nana, Anumeta henkei, Eupithecia cingulata и др. В июне еще продолжается лёт нескольких видов, но основной аспект фауны в это время, как и в последующие летние месяцы, составляют поливольтинные формы.
В особенностях лёта имагинальной фазы фактически отражается жизненный цикл данного вида, синхронно связанный с сезонным ритмом, в первую очередь, с климатическими условиями и развитием кормовых растений. Вместе с тем время появления и исчезновения имагинальной фазы, спаривание и откладка яиц в определенном сезоне года являются не только важным показателем для характеристики жизненного цикла, но и таким моментом в жизни насекомого, который в значительной степени определяет дальнейшее направление годичного развития.
Но рассмотрим несколько подробнее изменения в видовом составе весенних моновольтинных чешуекрылых, в основной своей массе представленных как раз весенне-раннелетними формами с преимагинальной диапаузой в период знойно-засушливого лета.
Вылупление бабочек приурочено к более или менее постоянным календарным срокам с некоторыми колебаниями по годам в зависимости от погодных условий. У ранневесенних видов, появляющихся в очень неустойчивое по температурному режиму время, эти колебания довольно большие. Разница во времени появления более поздних видов (поздней весной или ранним летом), в различные годы в соответствии с более постоянной погодой, оказывается значительно меньшей.
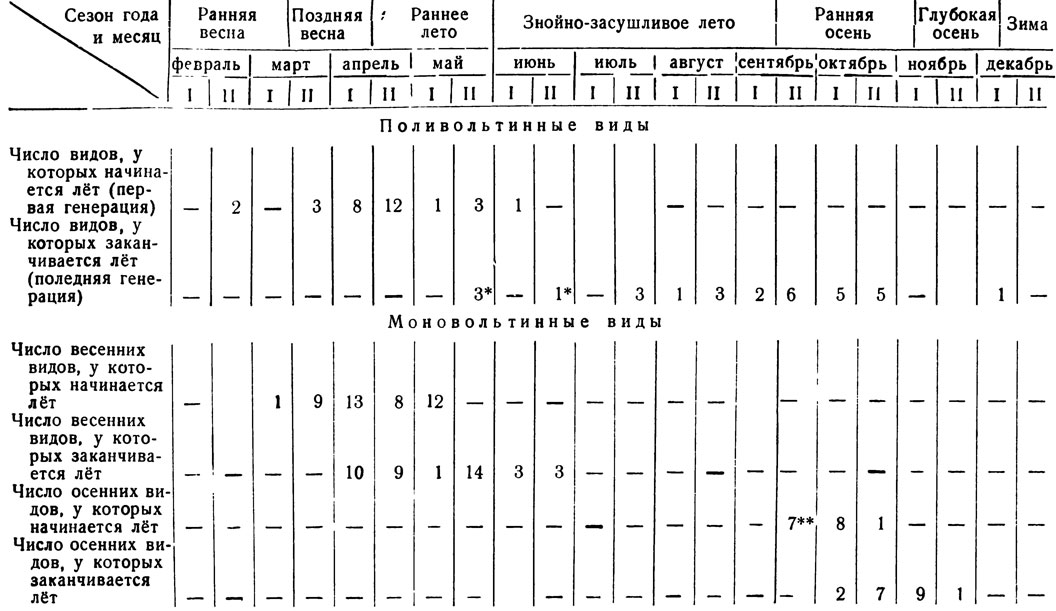
Начало вылета большинства весенних видов происходит со-второй половины марта до конца первой половины мая, т. е. с начала поздней весны до середины раннего лета (табл. 9). Наибольшее число форм появляется в первых половинах апреля и мая, когда среднесуточные температуры не падают ниже 10°С. Позже середины мая, когда среднесуточные температуры достигают примерно 23°С и поднимаются выше, к весенне-раннелетней фауне не добавляется ни одного моновольтинного вида, хотя лёт ранее появившихся форм еще продолжается. Ранней весной, в марте, отмечены только Lithostege luminosata Chr. и Euchloe belia pulverata Chr. Большинство видов заканчивает лет (табл. 9 и 13) в апреле (примерно поровну в первой и во второй половинах месяца - соответственно 10 и 9 видов) и во второй половине мая (14 видов). Последние три вида исчезают во второй половине июня. С конца июня в течение всего знойно-засушливого периода моновольтинные формы не встречаются, кроме Armada eylandti, запоздалые особи которой иногда попадаются в начале июля, и Satyrus enervata, вторичный лёт которой (после летней имагинальной диапаузы) начинается с первых чисел сентября, т. е. в конце знойного лета. Однако эта последняя принадлежит к следующей группе с иным типом годичного цикла, к рассмотрению которого и переходим.
Видов со вторым типом годичного цикла, летающих ранним летом и вторично осенью после летней имагинальной диапаузы, только 3: Satyrus enervata, Ogygia flammatra и Euxoa conspicua. Из них два последних - полигофаги с большими ареалами, охватывающими и значительную часть Европы. Все три формы связаны с травянистыми весенними растениями, но у S. enervata кормовыми растениями являются только злаки, и ареал ее в основном среднеазиатский. Окончательно не выяснено также, впадает ли имаго S. enervata в летнюю диапаузу, как два других вида (очевидно, именно так и обстоит дело), или на период летней жары бабочки мигрируют в горные районы, а к осени снова возвращаются в долины и низкогорья. Для всех видов характерно довольно позднее вылупление и последующий лёт бабочек, в основном во второй половине раннего лета и в начале знойно-засушливого сезона, т. е. когда среднесуточные температуры не спускаются ниже 18 - 23°С. Заканчивается лёт с наступлением сильной жары, в июне или в начале июля, когда бабочки впадают в летнюю диапаузу. Повторный лёт происходит ранней осенью (E. conspicua и О. flammatra), т. е. во второй половине сентября - в октябре, или с конца знойно-засушливого лета (с начала сентября), как это наблюдается у S. enervata. Биологически подобное поведение имагинальной фазы объясняется тем, что во время летней жары половые продукты не развиваются.* При вскрытии самок, вполне сформировавшиеся яйца обнаруживаются только с наступлением похолодания, в период осеннего лёта, когда среднесуточные температуры снижаются до 23 - 22°С. В это время происходит откладка яиц, которые зимуют. Гусеницы вылупляются рано весной, но вылет бабочек происходит, как сказано, сравнительно поздно, так как рассматриваемые виды отличаются относительно длительным периодом развития в фазах гусеницы и куколки.
* (Для E. conspicua, О. flammatra это установлено было еще М. А. Рябовым (1952) в условиях Дагестана и впоследствии подтверждено мною на таджикистанском материале (Щеткин, 1960), для S. enervata - моими наблюдениями.)
Из видов с третьим типом годичного цикла, лёт которых происходит ранним летом и вторично следующей весной (после длительной летне-зимней диапаузы имаго), в песках Вахшской долины отмечена только одна Autophila cerealis. Кладка яиц происходит ранней весной следующего года. Autophila cerealis экологически во многом сходна с видами предыдущей группы. Она встречается во всех биотопах долины, а также в горах, имеет довольно широкий ареал, захватывающий и Юго-Восточную Европу, Закавказье и страны восточного Средиземноморья. Вылупление бабочек происходит тоже довольно поздно, со второй половины раннего лета до начала знойно-засушливого сезона.
Четвертый тип годичного цикла составляют виды, летающие только ранней осенью, зимняя диапауза которых протекает в фазе яйца. Группа эта не многочисленна, всего 15 видов, но среди моновольтинных занимает по числу форм второе место, а в осенней фауне составляет основное ядро (табл. 10). Большинство видов с относительно небольшими ареалами. Преобладают пустынные формы, в том числе 6 псаммофильных (40% этой группы) и столько же эремофильных форм, но совсем нет эврибионтов. Здесь имеются монофаги и олигофаги, но много-ядных форм с сравнении с группой весенне-раннелетней относительно больше. Во всяком случае, из числа видов с более или менее известной биологией можно назвать следующих полифагов: Chondrostega pastrana, Lasiocampa eversmanni, Neoris stoliczkana, Sidemia apotheina и, вероятно, Agrotis lasserei.
Раннеосенние виды летают с середины сентября, когда среднесуточные температуры спускаются до 21°С и ниже и исчезают почти полностью в начале ноября при падении температур до 6 - 5°С. В этот же период происходит и повторный лёт (после летней имагинальной диапаузы) рассмотренных выше трех видов второй группы, а также единственного представителя пятой группы (см. ниже). Но этих видов всего лишь четыре, и они общую картину лёта осенних форм данной группы не нарушают, а наоборот, подчеркивают. Поэтому в табл. 9, характеризующей сезонные изменения видового состава, приведены данные по всем видам, летающим осенью (четвертой, второй и пятой групп).
Вылет из куколок почти всех осенних видов начинается во второй половине сентября или первой половине октября (примерно поровну). Во второй половине октября добавляется только один вид, зато у некоторых форм лёт заканчивается уже в первой половине октября. Большинство же видов исчезает в конце октября и в первой половине ноября (табл. 9).
Если общий лёт основной массы весенне-раннелетних форм растягивается на два с лишним месяца, то лёт большинства осенних видов в целом захватывает около полутора месяцев, т. е. протекает более "дружно" и в более сжатые сроки. Эта особенность в общем лёте осенних видов в сравнении с весенне-раннелетними выработалась исторически и биологически целесообразна, так как осенью погодные условия с каждым днем ухудшаются (температуры неуклонно снижаются, начинают выпадать осадки, растения заканчивают вегетацию), а весной и ранним летом происходят как раз обратные явления. Наибольшего видового разнообразия осенняя фауна достигает в октябре (в первой половине месяца 18 видов, во второй - 17) (табл. 10 и 13).
Календарные сроки окончания лёта осенних видов несколько колеблются по годам в зависимости от погодных условий. При этом отклонения в окончании лёта видов в различные годы, ввиду большей неустойчивости температурного режима, имеют больший размах, чем разница в календарных сроках появления бабочек.
Формы этой рассматриваемой группы откладывают яйца осенью, как правило, на растения уже прекратившие вегетацию (Neoris stoliczkana) или отмершие части однолетников (Lemonia tancrei, Lasiocampa eversmanni и другие, питающиеся весенними травянистыми растениями). Вероятно, этим и объясняется то обстоятельство, что среди осенних видов относительно больше многоядных форм (в сравнении с весенней группой). Вылупившиеся весной одноядные гусеницы имеют больше шансов погибнуть из-за отсутствия кормовых однолетних растений, так как последние могут и не вырасти в местах осенних кладок яиц. Развитие гусениц осенних видов начинается ранней весной с началом вегетации кормовых растений, среди которых у данной группы есть и эфемеры (преимущественно) и кустарники. Окукление происходит в конце весны или начале раннего лета. Для всех осенних форм характерна длительная летняя куколочная диапауза. Куколки покоятся либо в почве (у большинства видов), либо в коконах на стеблях растений. У последних поразительна способность к перенесению очень высоких дневных температур в раскаленной песчаной пустыне (например, у Eriogaster henkei). У большинства видов весенне-раннелетней группы летняя жара и засуха переживается также в фазе куколки, но куколка у них остается и на зимовку, до весны.
Наконец, к пятому типу циклов принадлежит лишь один вид Xylina exoleta, очень широко распространенный в Палеарктике, как на равнинах, так и в горах; полифаг. Бабочки, вылетая осенью, зимуют, весной вновь появляются и откладывают яйца. Большинство гусениц заканчивает развитие к началу мая и уходит в почву, где в земляном коконе пронимфа впадает до осени в диапаузу, более короткую в горах (в зависимости от высоты), но в Вахшской долине длящуюся 5,5 месяцев. С похолоданием, в первой половине (или начале второй) сентября, происходит окукление, а во второй половине октября - вылет бабочек.
Таким образом, в песках Вахшской долины высшие чешуекрылые по характеру лёта или, что тоже самое, по типу годичного цикла развития (так как последний наиболее ярко выражается именно в особенностях лёта и яйцекладки) составляют пять групп. Количественные соотношения групп с разными типами годичного цикла представлены в табл. 11. По числу видов господствуют моновольтинные весенние формы, на втором месте в этом отношении стоят поливольтинные формы без летней диапаузы, третье место занимают моновольтинные осенние виды. Форм с другими типами жизненного цикла в песках очень мало: все вместе они составляют лишь 10%. Для долины в целом с ее фауной, конечно, более разнообразной эта картина объемного соотношения групп с различными типами циклов остается в самом общем виде примерно такой же, но отклонения имеются; к тому же и число типов оказывается большим. Однако этот вопрос специально будет рассмотрен в последующих работах по Вахшской долине.
Таблица 11
Если теперь рассмотреть сезонную динамику видового состава в целом по отдельным фазам развития, то это будет выглядеть следующим образом. Имагинальная фаза чешуекрылых в песках Юго-Западного Таджикистана по количеству видов преобладает ранним летом, когда в течение этого сезона отмечается до 64 видов, т. е. 71 % от общего числа высших бабочек. В период с ранней весны до конца раннего лета встречается 74 вида, или 82,2% всей фауны высших. В период знойно-засушливого лета, когда некоторые моновольтинные формы еще встречаютсяв начале сезона или появляются в самом конце его, а все остальное время господствуют поливольтинные, отмечается только 35 видов. Почти столько же (36 видов) летает в течение периода с ранней до конца поздней, осени. Более подробно эти данные представлены в табл. 12. Однако, если рассмотреть распределение имагинальной фазы не по сезонам года, а по месяцам, то картина будет несколько иной. Из табл. 13 видно распределение бабочек по месяцам, как моно- и поливольтинных форм, так и всей фауны высших в целом. В последнем случае на кривой отчетливо выступают два пика: наибольшее число видов приходится на апрель - май до начала июня (с максимумом во второй половине мая), второй, меньший, пик приходится на сентябрь - октябрь с кульминационной точкой в первой половине октября. Эти данные будут особенно интересны в сравнении с другими биотопами долины, а также иных вертикальных поясов. Но это будет рассмотрено в последующих работах.
Таблица 12

Таблица 13
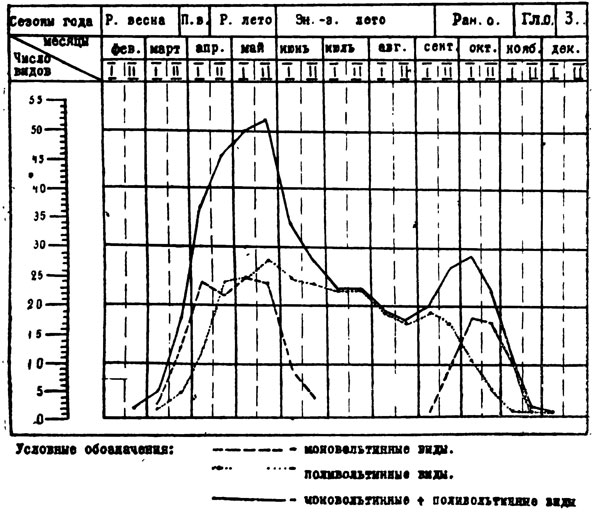
Фенология чешуекрылых в состоянии яйца остается недостаточно изученной. По примерным подсчетам эта фаза у преобладающего числа форм приходится на период раннего лета, когда происходит кладка у весенне-раннелетних моновольтинных и первых генераций поливольтинных видов, а также на сезон ранней осени, когда наблюдается откладка яиц осенних моновольтинных форм.
Личиночную фазу все без исключений моновольтинные виды проходят весной и ранним летом. В это время наблюдаются также гусеницы первого или второго поколений всех поливольтинных форм. В другие сезоны гусениц заметно меньше, а глубокой осенью и зимой они отмечаются лишь у 2 - 3 видов.
Наконец, фаза куколки преобладает в сезон знойно-засушливого лета, когда моновольтинные виды находятся в состоянии диапаузы, а также в начале ранней осени (до вылета осенних моновольтинных форм). Многие виды в фазе куколки переживают и суровые зимние условия.
Таким образом, приуроченность тех или иных фаз развития к определенным сезонам года находится в полном соответствии с климатическими режимами этих сезонов и генетическими особенностями видов, выражающимися, в частности, и в характере (типе) годичного цикла развития. Активные питающиеся фазы, гусеница и имаго, у большинства видов, как мы видели, приходятся на весну или раннее лето, когда пышно разрастается и цветет растительность. Лёт некоторой части бабочек приурочен к осени, когда цветущих растений вообще мало, а в песках они почти совсем отсутствуют. Но в песках у многих, если не у всех осенних форм имаго не питается совсем. Афагия отмечается у Chondrostega pastrana, Eriogaster henkei, Lasiocampa eversmanni, Lemonia tancrei, Neoris stoliczkana schencki, Agrotis lasserei unctus, Margelana versicolor, а также, вероятно, у Ochropleura danilevskyi, Scythocentropus scripturosa, Diadochia saca и Diadochia malitiosa usurialis.
На афагию многих осенних бабочек указывает также для кавказских подгрызающих совок и М. А. Рябов (1952). Можно добавить, что эта особенность осенних видов, пожалуй, еще в большей степени выражена у горных форм. В сезоны же, особенно неблагоприятные для жизни активных фаз, подавляющее большинство (в период знойного лета) или все виды (зимой) находятся на той или иной фазе (главным образом куколки или яйца) в состоянии диапаузы.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://butterflylib.ru/ 'Чешуекрылые, бабочки, мотыльки, моли'