Melanargia halimede Men. - Луговая пестрая бархатница и Melanargia epimede Stg. - Лесная пестрая бархатница
Эти виды систематически хотя и близки, но достаточно хорошо отличаются как по рисунку крыльев (см. определительную таблицу видов рода Melanargia), так и по строению половых органов самцов. Отличаются они по экологии и распространению. Если первый является жителем открытых травянистых биотопов (различные варианты разнотравных лугов и степи), то второй обитает в основном в редкостойных лесах (дубняки), но нередко и в других типах леса (долинные широколиственные и кедровники), встречаясь на полянах, по дорогам и берегам рек. Гусеницы М. halimede развиваются на вейнике (Calamagrostis epigeios), а М. epimede - на полевице (Agrostis clavata). Ареал луговой пестрой бархатницы более обширный. Она распространена по всему Приморью и по Амуру от г. Комсомольска до Зейско-Амурского плато включительно. Еще западнее, минуя наиболее холодные районы по Амуру и северные части гор Большого Хингана, она вновь начинает встречаться, но только спорадически по р. Шилке и в Забайкалье до г. Улан-Удэ. Кроме того, она распространена широко в Маньчжурии, в Восточной Монголии и на п-ове Корея.
Лесная пестрая бархатница имеет более узкий ареал. Кроме Приморья, она встречается в пределах среднего Приамурья и известна из Восточной Маньчжурии, Северного Китая и п-ова Корея.
Таким образом, эти виды на Дальнем Востоке по своему распространению дают типичный пример налегающих ареалов. Хотя морфологически и экологически они достаточно и обособлены, но, по моим наблюдениям, когда биотопы этих видов мозаично сочетаются, встречаются серии переходных форм между ними. Особенно явно и широко проявляются зоны интерградации между названными видами в районах сближения границ их ареалов (например, в бассейне нижнего течения рек - правых притоков Уссури и среднего Приамурья). Нередко отдельные участки переходных форм между М. halimede и М. epimede встречались и внутри ареалов (например, к западу и к югу от оз. Ханка в лесостепных и лугово-лесных районах). В связи с этим необходимо отметить, что целый ряд форм, относящихся к названным видам и описанных как раньше, так и за последние годы с п-ова Корея и прилегающих частей Китая (Wagener, 1959-1961) в качестве подвидов, с нашей точки зрения, являются мало или совершенно не обоснованными экологически и географически (например, М. h. chosenica Shelj., М. h. coreana, M. e. corimede Wag. и нек. др.) и скорее должны быть отнесены к гибридам двух разбираемых здесь видов рода Melanargia. По диагнозам и рисункам, приведенным в названной монографии С. Вагнера, некоторые из этих подвидов очень приближаются к нашим переходным формам между М. halimede и М. epimede, часто встречающимся в зонах интерградации этих видов.
В целом изучение в природе всех взаимоотношений между систематически близкими видами М. halimede, и М. epimede, которые раньше считались только подвидами (Seitz, 1932; Куренцов, 1957), даст многое для понимания процесса видообразования у чешуекрылых. Из своих наблюдений за этими видами я пришел к следующему заключению. М. halimede является более западным и сравнительно ксерофильным видом. В настоящее время происходит смещение его ареала к востоку и освоение им всех открытых ландшафтов. В результате сближения его ареала с ареалом М. epimede и возникли гибридогенные зоны между этими видами. Наблюдающиеся в этих зонах формы при условиях благоприятной эколого-географической изоляции могут вступить на путь таксономически более стойких образований. Допустить возникновение названных видов симпатрическим путем мы не имеем основания уже только потому, что ареалы этих видов имеют вполне определенные конфигурации и исторически, вероятно, сложились уже давно (рис. 50).
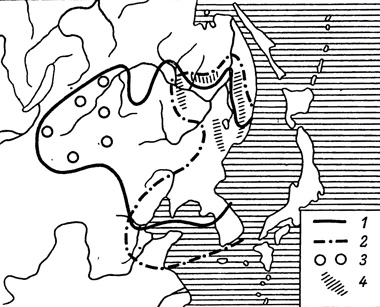
Рис. 50. Ареалы видов рода Melanargia. 1 - М. halimede; 2 - М. epimede; 3 - спорадические местонахождения М. halimede; 4 - зоны интерградации между этими видами
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://butterflylib.ru/ 'Чешуекрылые, бабочки, мотыльки, моли'